Вусачі
Вусачі́ (Cerambycidae) — п'ята за кількістю видів родина ряду Твердокрилих або Жуків, що належить до комах з повним перетворенням (голометаболією), які у життєвому циклі проходять стадії яйця, личинки, лялечки та імаго. За приблизними оцінками спеціалістів загальна чисельність родини становить приблизно 33 000 видів, приналежних до 5 200 родів, які розповсюджені по всьому світу[1], проте це далеко не повна оцінка, оскільки щорічно описують нові види з тропічних та екваторіальних регіонів, а іноді з Європи, Малої Азії й Північної Америки. В Україні описано близько 280 видів вусачів[2], для порівняння: в США — 1200, в Канаді — 350, а в Росії — 585[3][4]. Разом з тим вусачі є однією із найбільш вивчених родин, що зумовлено їхньою естетичною привабливістю й високою популярністю серед колекціонерів, зібрання яких мають ще й велике наукове значення, оскільки часто стають джерелом відкриття нових видів, використовуються науковцями-ентомологами при складанні фауністичних переліків на певних територіях тощо.
? Вусачі | ||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | ||||||||||||||||||||||||
| Біологічна класифікація | ||||||||||||||||||||||||
| ||||||||||||||||||||||||
| ||||||||||||||||||||||||
Посилання | ||||||||||||||||||||||||
| ||||||||||||||||||||||||
Сім видів вусачів занесено до Червоної книги України.
Вагомий внесок у вивчення цієї родини внесли українські вчені Д. В. Зайців, І. К. Загайкевич, Д. Ф. Руднєв та О. Ф. Бартєнєв.
Назва
Назва родини «Cerambycidae» є латинським неологізмом, який не існував у класичній латині, а був створений у часи Карла Ліннея. Вона сформована зіставленням двох слів: давньогрецького ceros — «ріг» та латинського bis — «два», що означає «дворогий»[5]. Ця назва відображає особливості їх антен — довжину та їх прижиттєве спрямування вперед, що асоціюється з рогами. У багатьох західних і центрально-європейських мовах назва вусачів пов'язана саме із рогами: англ. Longhorn beetle, фр. Longicornes, ісп. Escarabajos longicornios, нім. Bockkäfer, пол. Kózkowate тощо. У той же час у південних та східно-європейських мовах закріпились дві відмінні одна від іншої назви. Перша асоціює антени із вусами: укр. Вусачі, рос. Усачи, біл. Вусачы тощо. Друга — із живленням деревиною їх личинкової стадії: рос. Дровосеки, біл. Дрывасекі, болг. Сечковци, чеськ. Tesaříkovití тощо[6].
В українській мові, окрім загальноприйнятої та основної назви «Вусачі», відомі також такі: Скрипуни́, Скрипунува́ті, Довгоро́жці, Ковзани́́ та Козаки́[7]. Назви «Скрипуни» та «Скрипунуваті» пов'язані із особливостями їхньої поведінки — імаґо вусачів, раптово схоплені, видають скриплячий звук, тручи передньоспинкою об середньоспинкиу; назва «Довгорожці» вказує на подібність антен до рогів; «Ковзани» — на подібність антен на ковзани (полози) сан; «Козаки» — на подібність антен на довгі вуса козаків.
Морфологія
Імаго
Тіло дорослих жуків або імаго, як і решти комах, складається з трьох основних відділів: голови, грудей та черевця.

%252C01.JPG.webp)

Голова вусачів має округлу форму й своєю задньою частиною входить у передньогруди. В межах родини розрізняють два типи голови: перший — це спрямована вперед або косо донизу під невеликим кутом до центральної осі тіла (Прионіни, Лептурини, Церамбіціни, Асеміни, Нецідаліни), друга — спрямована вниз під кутом 90° до основної осі тіла (Ляміїни). На боках голови розміщені складні, багатофасеткові очі, які мають типову для вусачів ниркоподібно- виїмчасту форму. Іноді очі цілком розділяються на дві частини — складається враження, що в жука дві пари очей (декотрі Ляміїни). На верхній стороні голови розрізняють кілька слабко розмежованих частин: шию, потилицю, скроні, тім'я, чоло та наличник. Шиєю називається та частина головної капсули, яка входить у груди. Та частина голови, що безпосередньо примикає до шиї, називається потилицею і має випуклу форму. По обидві сторони від потилиці у вусачів наявні характерні виступи — скроні. Між очима, вище потилиці розміщене тім'я, яке на передній частині переходить у лоб.
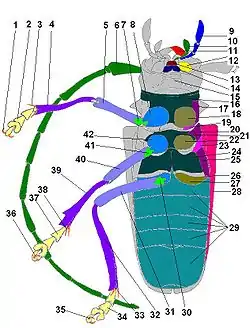
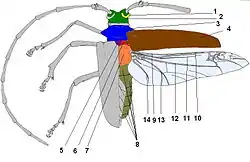


На лобі, між очима, розташовані вусики (антени), основи яких містяться у вусикових впадинах. Вусики складаються з 11-ти члеників, а в деяких родів (Agapanthia, Phytoecia, Stenostola тощо) — з 12-ох. У представників родів Monochamus, Lamia, Morimus перший членик має цикатрикс — характерний півмісячний плоский виступ поруч з вершиною. Вусики здебільшого перевищують довжину половини тіла, щетинкоподібної або нитчастої, зрідка пильчастої або гребінчастої форми і за життя закидаються на спину, й можуть вкладатися тільки вздовж верхньої сторони тіла — це одна із ознак, що відрізняє родину Вусачів від інших родин Твердокрилих. Довжина вусиків самців значно перевищує їх довжину у самок. На передній частині лобу наявний шов, який відмежовує від нього наличник. З боків голови, нижче очей, розміщені щоки. До наличника причленовується верхня губа, яка частково прикриває мандибули. На нижній частині голови між шиєю та ротовим апаратом знаходиться горло. Його передня і найвужча частина, яка прилягає до нижньої губи, називається субментумом або підборідком, а поздовжні боріздки біля його основи — горловими швами. Вусачі мають гризучий ротовий апарат, який складається з мандибул або щелеп, верхньої губи, нижньої губи, підборіддя, губних та щелепних щупалець.

Груди складаються з трьох сегментів, з яких найрозвиненішим є перший. З дорзальної сторони вони називаються спинками: передньо-, середньо- та задньоспинкою.
Пердньоспинка часто має боковий шип (у вусача ялинового, дубового, вусача-товсуна, вусача-раґія тощо) або горбик (у вусача-карілія, пахіти тощо), а у інших просто заокруглена (наприклад, в анастранґалії, лептуробоски, кліта-імітатора, плосковусача). Зверху помітною є лише пердньоспинка, а середньоспинка, окрім щитка, та задньоспинка приховані надкрилами. Надкрила кріпляться до середньоспинки, а до задньоспинки — перетинчасті крила, призначені для польоту. Декотрі види, як от вусач-товстун, морімус, вусач-коренеїд, втратили здатність до польоту і крила у них редуковані або відсутні цілком. Частина надкрил, яка заходить на нижню сторону тіла, називається епіплеврами. Майже, всі види вусачів здатні видавати скрипучі звуки за рахунок тертя виросту середньоспинки до заднього краю передньоспинки. З нижньої сторони груди складаються з передньо-, середньо- та задньогрудей. Кожен з цих сегментів, в свою чергу, складається з центральної частини — диску, відмежованих від нього бічної — епістерн, та задньої — епімер, сторін. В задній частині кожного сегменту грудей знаходяться тазикові впадини, в яких розташовані, власне, тазики — частина кінцівки.

Кінцівки складаються з п'яти основних частин: тазиків, вертлюгів, стегон, гомілок та члеників лапок. У вусачів передні тазики мають кулеподібну або конусоподібну форми, середні, за звичай, є кулястими, а задні — поперечні. Тазик зі стегном сполучає вертлюг, з яким він вільно рухається. До вершини стегна приєднується гомілка, а до неї лапка з члениками. На вершині гомілнки розташовані дві шпори, у деяких видів — по одній. Лапка має вигляд чотиричленикової, оскільки, власне, четвертий членик є дуже малим і, практично, непомітним. Третій членик розділений на дві частини і є дволопатевим. Останній членик лапки несе два кігтики. Усі членики лапок більш-менш розширені з густою щіточкою щетинок на нижній стороні.
Черевце складається з п'яти вільних кілець. З верхньої сторони, прикритої надкрилами, вони називаються тергітами, а з нижньої — стернітами. По краю тергітів розташовані дихальця, через які жуки дихають. Останній членик черевця називається пігідієм.
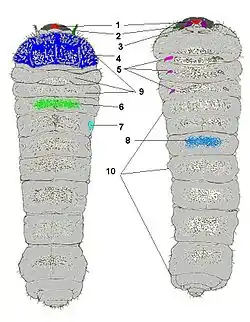
Личинка
Тіло личинок сплющене або більш-менш призматичної форми, м'ясисте, білого чи жовтуватого кольору з темнішою передньоспинкою та головою, остання втягнена в груди зі спрямованими вперед щелепами. На голові розміщені дуже малі вусики — тричленикові, та очі. В еволюційно розвинених груп вусачів очі рудиментарні або відсутні взагалі (роди Кліт, Аґапантія, Фітеція), тоді як у примітивніших, окрім основних очей, наявні ще й додаткові вічка (роди Вусач-шкіряник, Спондиліс, Плосковусач тощо). Очі у житті личинки не відіграють жодної ролі, адже вона живе усередині субстрату (деревини, стебел рослин чи ґрунту), куди світло не проникає. На верхній стороні сегментів тіла наявні мозолисті горбики по одному на кожному сегменті та по два овальні дихальця. Ніжки короткі ледь помітні. У еволюційно примітивних таксонів вусачів (роди Prionus, Rhagium, Gnathacmaeops тощо), на кінчику черевця личинки, наявні гачечки або шипи, які допомагають їм переміщуватись у вигризених тунелях у зворотному напрямку. Личинки ж високоорганізованих родів (Phytoecia, Agapanthia тощо) таке пристосування втратили.
Лялечка
Вусачам притаманний відкритий тип лялечок — зачатки крил, кінцівок і вусиків є вільними (не зрощеними), лише притиснуті до тіла. Заляльковування у різних таксономічних груп має свої відмінності. У більшості Прионін, а також декотрих примітивних Лептурин із триби Раґіїні (Вусачик-карілія, Пахіта тощо), личинка останнього віку виходить з деревини у ґрунт, де будує лялечкову камеру й заляльковується. Схожа поведінка притаманна і Доркадіїнам (підродина Ляміїни), проте їхні личинки живуть у ґрунті, де і заляльковуються. Більшість Церамбіцін, Лептурин, Ляміїн та Асемін зяляльковується у деревині — декотрі вгризаються у товщу дерева, де зі стружок будують лялечкову камеру (наприклад, вусач дубовий, вусач ялиновий, вусач хатній), інші заляльковуються під корою (наприклад, Вусач-раґій). Прогресивні триби Ляміїн — Аґапантіїні, Фітеціїні тощо — заляльковуються у стеблах та кореневищах різноманітних трав.
Життєві цикли




Більшість вусачів мають одно- або дворічний цикл розвитку, який представлений повним перетворенням або метаморфозом. У різних груп вусачів немає принципових відмінностей у їхньому розвитку, усі вони проходять стадії яйця, личинки, лялечки та імаго. Відмінності існують лише у шлюбній поведінці та способах відкладання яєць, проте існує кілька екологічно відмінних життєвих циклів у вусачів. Перший — це розвиток личинок у деревині з додатковим живленням на стадії імаго пилком квітів — антофіли (роди Nivellia, Carilia, Leptura тощо), вегетативними органами дерев та їх соком (Monochamus, Acanthocinus, Cerambyx тощо) або взагалі без додаткового живлення — афаги (Prionus, Hylotrupes тощо). Другий — це розвиток личинкової стадії у стеблах трав'янистих рослин з родин Айстроцвітих й Зонтичних з додатковим живленням на імагінальній стадії вегетативними частинами трав'янистих рослин (вусачі з родів Agapanthia, Phytoecia тощо). Третій — це розвиток личинкових стадій у ґрунті з їх живленням коренями злаків, а імагінальні форми додатково живляться їх наземними частинами (наприклад, вусачі Dorcadion).
В першому варіанті життєвого циклу самка групами відкладає яйця на поверхню або у тріщини кори дерева. Така кладка притаманна більшості представників підродин Лептурини, Прионіни та Церамбіціни, що вважається дуже примітивною ознакою, адже відсутня будь-яка турбота про потомство. У видів з підродини Ляміїни процес відкладання яєць дещо складніший. У найпримітивнішому варіанті самка, наприклад, вусача ялинового великого західного вигризає на корі дерева спеціальну ямку — насічку, куди опускає яйцеклад і відкладає одне яйце. Значно складніша поведінка у скрипуна осикового (Saperda populnea) — самка на тоненьких гілочках осики робить насічку, куди відкладає одне яйце й прикриває його вигризеною нею стружкою з кори. Після того самка по колу обгризає верхівку пагона так, що через кілька днів вона в'яне, і в мить виходу личинки з яйця всихає та відпадає. Такі дії самки призводять до ослаблення пагона, в якому розвиватиметься личинка, і перешкоджають рослині виділяти захисні речовини — терпени. Личинки після виходу з яйця живляться лубом. Личинка Saperda populnea відрізняється від більшості вусачів своєрідним способом життя. Вона, спочатку, харчується новоутвореною тканиною біля насічки, яку підготувала самка, потім робить вузький кільцевий периферійний хід в лубі. До осені, в більшості випадків, заглиблюється в серцевину, де прокладає вгору короткий (близько 4 см) вертикальний хід. Дерево реагує на пошкодження утворенням «пухлини». У Прионін і Лептурин личинки розвиваються у мертвій деревині, яка почала гнити, заселена бактеріями та грибами. Личинки більшості видів Лептурин пов'язані з грибами, сумішшю міцелію яких і деревини вони живляться.
У видів, що мають дворічну генерацію, личинка наприкінці літа вгризається у глибину деревини, де зимує, а навесні повертається під кору, де продовжує живитись і рости, заляльковуючись лише восени. У вусачів з однорічним життєвим циклом, личинка наприкінці літа також вгризається у глибину деревини, де вигризає лялечкову камеру та заляльковується. Навесні або влітку наступного року дорослий жук проходить через личинковий тунель, прогризає кору дерева та виходить назовні. У видів, що розвиваються у коренях дерев, личинки заляльковуються у ґрунті, наприклад, Prionus, Pachyta, Carilia тощо. Личинка вусача великого дубового західного живе під корою, де, живлячись, вигризає довгі звивисті ходи. Перед заляльковуванням вона вгризається в деревину, прокладаючи короткий гачкуватий хід, який завершується лялечковою камерою. В цій камері личинка створює своєрідну стінку з дрібної тирси, яка закриває личинковий хід. Така стінка забезпечує стабільний температурний режим в камері та захищає від зовнішніх ворогів, які могли б потрапити в личинковий хід. Лялечка формується восени або в кінці літа й жук зимує в лялечковій камері, виходячи на поверхню лише наступного року, на початку літа. Він спершу прогризає отвір в личинковий хід, а від так прогризає кору й виходить назовні.
Походження


Жуки-вусачі є відносно давньою родиною. Якщо перші палеонтологічні дані щодо жуків взагалі стосуються пермського періоду (250 млн років тому) — кінця палеозойської ери[8], то представники родини Вусачів у викопному стані відомі лише з юрської системи (130 млн років тому) — середини мезозойської ери[3]. З цього періоду описані як вимерлі, так і деякі сучасні таксони вусачів. Наприклад, Prionus ooliticus в лейасових відкладах; Cerambycinus dubius Germar із Золенгофенського літографічного сланцю[3][9]. Таким чином у верхній юрі розвинулись первісні форми вусачів, з яких сформувалася власне родина. Вони мали багато спільних рис з листоїдами, які також зароджувались у цей час. В нижній крейді первісні форми вусачів досягають значного різноманіття й цілковито відокремлюються від листоїдів, що призводить до початку формування вищих таксонів родини, а уже до середини крейдового періоду вони вимирають. Їх місце займають представники давніх підродин прионін, парандрин, лептурин та інших, що припадає на час появи перших покритонасінних, деревину яких вони заселяють. Одночасно від гілки еволюції вусачів відокремлюються родини Веспериди та Дистениїди, які з початком кайнозойської ери досягали значного видового різноманіття, а з початком зледенінь вимерли. В третинному періоді вусачі захоплюють практично всі екологічні ніші, які існують в умовах тліючої деревини й починають заселяти живі дерева, набуваючи високого рівня видового різноманіття. В третинних відкладах зустрічаються здебільшого сучасні види. Так, із буровугільної формації описано види родів Leptura, Lamia, Astynomus й інші, та знайдені як самі жуки так і ходи їх личинок в деревині — Astynomus tertiarius. Із середини третинного періоду з'являються трав'янисті покритонасінні рослини, що стає поштовхом до розвитку багатьох нових таксонів вусачів таких, як Dorcadiini, Agapanthiini, Phytoeciini. Ці види переходять до життя в стебла рослин або ґрунт. Середина третинного періоду є часом розквіту дводольних — значного поширення набувають Зонтичні, Шорстколисті, Айстроцвіті та інші рослини, у стеблах яких поселяються вусачі. Початок четвертинного періоду та зледеніння спричинили вимирання значної частини видів, поширених у помірних зонах обох півкуль. У Європі та Північній Азії зникли третинні широколистяні ліси, що зумовило вимирання більшості тутешніх вусачів. Третинна реліктова флора та фауна цих лісів збереглася лише на Далекому Сході й характеризується значним видовим різноманіттям[10].
Філогенія
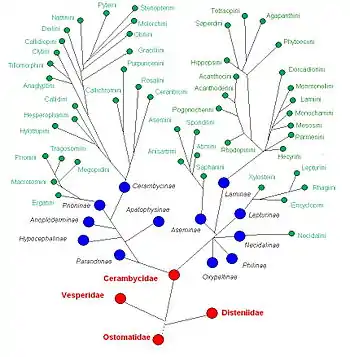
Філогенія родини вусачів є складним питанням, яке викликає дуже багато дискусій у середовищі спеціалістів, що обумовлено великою кількістю видів та їх широким розповсюдженням. Вусачі — центральна родина надродини Церамбікоїдних, яка була виділена П'єром Латреєм у 1802 році, проте згодом віднесена до надродини Хризомелоїдних, з якою вони мають низку спільних морфологічних ознак і є найближчими родичами. Обидві ці надродини мають спільного предка або походять від надродини Кукуїдоїдних жуків[3]. Надродина Церамбікоїдні утворює дуже компактну та морфо-екологічно добре окреслену групу, що включає чотири родини, — окрім Вусачів, сюди також належать Веспериди, Дистениїди та Оксипельтиди.
Згідно із сучасними уявленнями родина Вусачів включає дві більш-менш філогенетично віддалені гілки[11]. Перша — це підродини Прионіни та Церамбіціни, які походять від спільного предка, причому підродина Прионіни є більш примітивною й реліктовою групою вусачів. Як вважалось раніше[3], разом із цією гілкою розвинулись підродини Парандрини, Апатофізини, Анопльодерміни та Гіпоцефаліни. На сьогодні підродину Парандрини розглядають як трибу Парандріні у складі підродини Прионіни. Підродину Апатофізини переведено до рангу триби Апатофізіні і зведено у синоніми до Доркасоміні (Dorsacomini) у підродині Лептурини. Аноплодерміни та Гіпоцефаліни переведені до родини Веспериди. Часом Аноплодермін виділяють в окрему родину[12]
Друга філогенетична гілка родини Вусачів включає центральний стрижень Лептурини — Ляміїни, разом з ними розвинулись невеличкі підродини Нецідаліни та Асеміни. Раніше до цієї групи також відносили підродини Оксипельтини та Філіни[3], нині, першу виділяють як окрему родину Оксипельтиди, а другу переведено до родини Дистениїди[13].
Систематика
Родину вусачів, за різними системами, розділяють на 7—10 підродин. Сім з них відносять до вусачів (наведені нижче), а три виокремлюють в ранг родин (дивіться статтю Церамбікоїдні):
- Парандрини (Parandrinae Latreille, 1802)
- Прионіни (Prioninae Latreille, 1802)
- Лептурини (Lepturinae Latreille, 1802) — також виокремлюють Апатофізини
- Нецідаліни (Necydalinae (Latreille) Pascoe, 1864)
- Асеміни (Aseminae Thomson, 1860) — також виокремлюють Спондиліни
- Церамбіціни (Cerambycinae Latreille, 1802)
- Ляміїни (Lamiinae Latreille, 1802)
Роль в екосистемах




Роль вусачів в природних екосистемах полягає в тому, що, будучи консументами першого порядку, вони є невід'ємними ланками ланцюгів харчування, зв'язковими між окремими складниками лісових екосистем[14]. В процесі коеволюції організмів сформувалась харчова мережа зв'язків між автотрофами та гетеротрофами, в яку вусачі включені на стадіях личинки, лялечки та імаго. Оскільки личинки та лялечки розвиваються в такому консервативному середовищі як деревина, де умови розвитку не мінялися протягом десятків та сотень мільйонів років, то вони є ланками в живленні птахів й паразитичних комах-ентомофагів. Імаго включаються в трофічну мережу значно ширшого спектру консументів другого порядку — це різноманітні птахи, комахи-хижаки, дрібні ссавці тощо.
Консортивні зв'язки з деревними рослинами
Як комахи-ксилофаги личинки більшості видів вусачів, за деякими винятками, розвиваються в деревині — вони є утилізаторами деревного відпаду хвойних та листяних порід дерев. Жуки-вусачі, в значній мірі опосередковано, беруть участь у мінеральному живленні рослин, яке виражається в поверхневому надходженні зольних елементів і азоту із зоогенним опадом, екскрементами та стимуляцією деструктивних процесів. По суті, вусачі разом з деревними рослинами, в відпаді деревини яких вони розвиваються, утворюють популяційні консорції, де їхня функціональна роль полягає у звільнені шляхом деструкції мертвої органіки елементів, які того ж року будуть використані для формування живої органічної маси детермінанта консорції[15].
Шкодочинність
Вусачів відносять до одних з найсерйозніших шкідників деревних будматеріалів[16]. Вусачів, за шкодочинністю, поділяють на фізіологічно-технічних шкідників — види, що ушкоджують живі та ослаблені дерева і технічних шкідників — види, личинки яких заселяють уже мертву деревину[10]. Загалом, личинка у вусачів є основною шкодочинною стадією у життєвому циклі комахи[17].
Класичним прикладом шкодочинності вусачів є масове розмноження вусача дубового великого західного в 1909—1930 роках у Центральній та Північній Україні[18]. З 1872 до 1930 року на цих теренах було запроваджено так звані посмужні рубки лісу з шириною смуги від 40 до 80 м, окрім того на лісосіках залишали поодинокі дуби для вирощування «великомірного» лісоматеріалу або насінників. Посмужні лісосіки призвели до значного ослаблення та пригнічення життєвості як поодиноких дубів на зрубах, так і деревостанів, що безпосередньо примикали до вирубок, внаслідок зміни мікрокліматичних умов. Мікроклімат під шатром лісу характеризується більш сталою вологістю та меншими амплітудами добових температур, ніж на відкритій місцевості. Ослаблені дерева ставали привабливими для заселення вусачами, що й спричинило спалах чисельності виду. Заходи боротьби у таких випадках малодієві. На сьогодні великий дубовий вусач вважається рідкісним видом і занесений до Червоних списків та книг всіх європейських країн, в тому числі й України.
Ще одним хрестоматійним прикладом є спалах чисельності вусача ялинового Розенмюллера (Monochamus rosenmuelleri) в Томській області та Красноярському краї СРСР в 1961 році[19]. Внаслідок масштабного вирубування лісу на площі близько 50 000 га спостерігали масове розмноження цього вусача, зумовлене неправильним господарюванням. Одним із заходів вирішення цієї проблеми було прискорення вирубувань та збільшення їхніх площ. Така діяльність призвела до швидкого зростання чисельності цього виду й загибелі лісів на широких просторах сибірської тайги. Характерними ознаками була поява значних за площею ушкоджених ділянок лісу, на яких спостерігалось пожовкнення й осипання хвої сибірської модрини, сибірської ялини та сибірської ялиці, такі ділянки не придатні для промислового використання. Як зауважує А. Рожков, у даному районі спостерігали масові скупчення комах, які повністю обгризали луб і хвою дерев, ослаблюючи їх. Окрім того дорослі комахи відкладали яйця в ослаблені дерева, а розвиток личинки спричинював загибель останніх. Ефект масового розмноження великого ялинового вусача в цьому регіоні спостерігався впродовж 10 років, поки не було припинено будь-які вирубування лісу.
З метою боротьби із вусачами-шкідниками господарських рослин та лісового господарства, використовують як класичну фумігацію інсектицидами, так і біологічні, а також біохімічно-фізіологічні способи. Одночасно, розробка таких методів надає матеріал для наукових досліджень фундаментальнішого характеру. Зокрема, для розробки методів боротьби з жуком Xylotrechus pyrrhoderus, що є одним з найнебезпечніших шкідників винограду у Південно-Східній Азії, була виділена та переведена до сталого автономного існування in vitro клітинна лінія XP-1 з клітин його жирового тіла, на якій потім тестувалася цитологічна чутливість до кількох гормонів.[20] Ще одна стабільна клітинна лінія вусачів, РС-1, була отримана японськими дослідниками з жука Plagionotus christophi в 2008 році[21].
Санітарна роль у лісових екосистемах
В природних і незначною мірою змінених екосистемах, вусачі виконують санітарну роль, елімінуючи з деревостанів та утилізуючи ослаблені й пригнічені конкурентною боротьбою за світло, суховершинні, вражені кореневою губкою (Heterobasidion annosum (Fr.) Bref.), трутовиками (Phomitopsis pinicola (Sw. ex Fr.) Karst., Ph. chrysoloma (Fr.) Donk., Polystictus circinatus (Pers.) Bres. та ін.), кореневими гнилями дерева, до певної міри регулюючи поширення цих грибів. Особливу роль вони відіграють на вітровалах та зрубах, де прискорюють розклад мертвої деревини[22]. Елімінація таких дерев звільняє місце для підросту й сприяє відновленню екосистеми.
Запилення квіткових рослин
Вусачі також виступають запилювачами багатьох видів квіткових рослин, адже більшості цих комах притаманна антофілія. Зокрема антофільними є більша частина видів підродини Лептурини, а дещо менше види з підродини Церамбіціни. Імаго цих видів трапляються на квітах, де на відміну від «класичних» запилювачів (бджіл, джмелів, метеликів, дзюрчалок тощо) вони проводять значно більше часу, що підвищує ефективність запилення. Вусачі регулюють чисельність запилюваних ними квіткових рослин. Це пов'язано з тим, що дорослі комахи для повного статевого дозрівання та дозрівання яєць харчуються пилком і часто-густо поїдають гінецей та андроцей, зменшуючи насіннєву продуктивність рослин. Як правило, рослини, які запилюються вусачами, мають дуже високу насіннєву продукцію й поїдання їх генеративних органів не спричинює особливих втрат.
Див. також
Примітки
- Adam Slipinski, Hermes Escalona (2013) Australian Longhorn Beetles (Coleoptera: Cerambycidae) Volume 1: Introduction and Subfamily Lamiinae (англ.)
- Бартенев А. Ф. Обзор видов жуков-усачей (Coleoptera: Cerambycidae) фауны Украины // Вісник Харківського ентомологічного товариства. — 2003 (2004). — 11, № 1-2. — с. 24-43
-
- Загайкевич И. К. . Таксономия и экология усачей. — К.: Наукова Думка, 1991. — 420 с.
- Danilevsky M.L. Taxonomic and zoogeographical notes on the family Cerambycidae (Coleoptera) of Russia and adjacent regions // Russian entomological journal. — Moscow, 1993. — vol. 1, No. 2. — pp. 37-39;
- Hildt L. F. 1917. Owady krajowe Kózkowate. Cerambycidae. Pam. Fizyogr., Warszawa, 24 (III), str. 1-141, 6 pl.
- Жуки-вусачі Українських Карпат. Розмаїття.
- Словник зоологічної номенклатури (1927—1928 рр) — К.: Наукова думка, 2005. — 186
- Ponomarenko A.G. The geological history of beetles // in: J. Pakaluk & S.A. S'lipin'ski (eds.). Biology, Phylogeny, and Classification of Coleoptera. Papers celebrating the 80th Birthday of Roy A. Crowson. — Warszawa: Muz. Inst. Zool. PAN — 1995 — Vol. 1 — P. 155—172.
- Barthel K. W., Swinburne N. H. M., Conway Morris S. Solnhofen. A Study in Mesozoic palaeontology. — Cambridge University Press, 1990. — P. 151. — ISBN 9780521333443.
- Шаблиовский В. В. Жуки-дровосеки лесной зоны СССР // Автореф. дисс. д-ра биол. Наук: 03.00.09. — Л., 1967. — 44 с. (рос.)
- Napp D. S. 1994 — Phylogenetic relationships among the subfamilies of Cerambycidae (Coleoptera, Chrysomelidea) — Revista Brasileira de Entomologia 28 (2): 265—419
- Jenis I., 2001 — Long-Horned Beetles Distenidae, Oxypeltidae, Vesperidae, Anoplodermatidae & Cerambycidae I, Vesperidae and Cerambycidae of Europe I, A. Regulus, Zlin, 333 pp.
- Švacha P., Wang J. J. & Chen S. C. Larval morphology and biology of Philus antennatus and Heterophilus punctulatus, and systematic position of the Philinae (Coleoptera: Cerambycidae and Vesperidae). Annales de la Societé entomologique de France (N. S.). — 1997. — 33 (3): 323—369.
- Заморока А. М., Парпан В. І. Сезонні флуктуації видового багатства жуків-вусачів (Coleoptera: Cerambycidae) в лісових екосистемах північно-східного макросхилу Українських Карпат та Передкарпаття // Питання біоіндикації та екології. — Запоріжжя: ЗНУ, 2007. — вип. 12, № 1. — с.125-133
- Царик Й. В., Царик І. Й. Консорція як один із базових рівнів біологічного різноманіття // Карпатський регіон і проблеми сталого розвитку: Матеріали конф. — Рахів, 1998. — с. 303—304
- Шаблиовский В. В. Жуки-дровосеки Приморского края // Автореф. дис. канд. сельскохозяйственных наук: 03.00.09. — Л., 1950. — 15 с.
- Данилевский М. Л. Морфо-экологические закономерности эволюции личинок жуков-дровосеков // Автореф. дисс… канд. биол. наук: 03.00.09. — М., 1976. — 17 с. (рос.)
- Руднєв Д. Ф. Великий дубовий скрипун (Cerambyx cerdo L.). — К.: видавництво Всеукраїнської Академії Наук, 1935. — 141 с.
- Рожков А. А. Про большого елового усача // Химия и жизнь. — 1976. — № 12. — с.88-92
- K. Iwabuchi. An Established Cell Line from the Beetle, Xylotrechus Pyrrhoderus (Coleoptera: Cerambycidae)[недоступне посилання з листопадаа 2019] // In Vitro Cell. Dev. Biol.-Animal. — 1999. — Vol.35, — pp. 612—615
- K. Hoshino & M. Hirose & K. Iwabuchi. A new insect cell line from the longicorn beetle Plagionotus christophi (Coleoptera: Cerambycidae)[недоступне посилання з листопадаа 2019] // In Vitro Cell. Dev. Biol.-Animal. — 2008. DOI:10.1007/s11626-008-9152-7
- Wermelinger B., Duelli P. Obrist M. K. Dynamics of saproxylic beetles (Coleoptera) in windthrow areas in alpine spruce forests // For. Snow Landsc. Res. — 2002. — Vol. 77, ½. — pp. 133—148
Література
- Екологічні особливості ентомокомплексів жуків-вусачів (Coleoptera: Cerambycidae) у лісових екосистемах північно-східного макросхилу Українських Карпат: Дис. … канд. біол. наук: 03.00.16 / А. М. Заморока. — Івано-Франківськ, 2009. — 209 c.
Посилання
- Вусачі Галичини та Карпат у базі даних Fauna Haliciae et Carptiae (укр.)
- Систематика родини на порталі BioLib (англ.)
- Фотогалерея вусачів світу (англ.)
- Вусачі західної палеарктики (англ.)
- Личинки та лялечки вусачів (пол.)
- Вусачі Фінляндії та Скандинавії (англ.)
- Вусачі Флориди (англ.)
- Вусачі світу (англ.)
- Вусачі Французької Ґвіани (фр.)
- Фотогалерея Станіслава Крейчика[недоступне посилання з червня 2019] (англ.)
- Вусачі палеарктики (англ.)
- Фотогалерея вусачів Яна Грбека (чеськ.)
|
|
Ця стаття належить до вибраних статей Української Вікіпедії. |