Твердокрилі
Твердокри́лі, або жуки́ (Coleoptera Linnaeus, 1758) — комахи з повним перетворенням, які у життєвому циклі проходять стадії яйця, личинки, лялечки та імаго. Свою назву — твердокрилі, жуки отримали у зв'язку із перетворенням передньої пари крил в хітинізовані тверді надкрила, які використовуються в польоті за принципом крила літака. Жуки — це одна із найбільших груп комах і живих істот взагалі — ряд нараховує понад 400000 видів. Нині прийнято вважати, що кожна п'ята тварина на Землі — це жук. Розміри коливаються в широких межах — від 0,3 мм у жуків-пір'єкрилок до 150 мм у Жука-Геркулеса чи 170 мм у Вусача-титана гігантського.
? Твердокрилі | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | ||||||||||||||||
| Біологічна класифікація | ||||||||||||||||
| ||||||||||||||||
Підряди | ||||||||||||||||
Посилання | ||||||||||||||||
| ||||||||||||||||
60 % всього різноманіття жуків складають представники лише п'яти родин: Жуки-слоники (Curculionidae) — 60 тис. видів, Жуки-хижаки (Staphylinidae) — 58 тис., Туруни (Carabidae) — 40 тис., Листоїди (Chrysomelidae) — 35 тис. та Вусачі (Cerambycidae) — 35 тис. 44 види твердокрилих занесено до Червоної книги України.
У фауні України кількість видів жуків досі остаточно не визначено. Вважають, що їх налічується не менше, ніж 25—30 тисяч видів[1]. Серед відомих українських колеоптерологів — К. Гормузакі, К. А. Пенеке, С. І. Медведєв, Д. О. Оглоблін, Д. В. Знойко, Ф. К. Лукьянович, В. Г. Долін та інші. Вагомий внесок у вивчення колеоптерофауни України зробив В. Г. Плігінський.
Морфологія
Тіло дорослих жуків або імаго, як і всіх інших комах, складається з трьох основних відділів: голови, грудей і черевця.
Будова голови

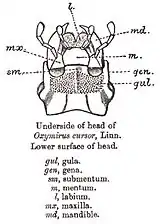
Голова жука вкрита суцільною капсулою, яка в ході еволюції, утворилася в результаті злиття кількох сегментів тіла та акрона, і має округлу форму. Своєю задньою частиною голова входить у передньогруди. На верхній стороні голови розрізняють кілька слабко розмежованих частин: шию, потилицю, скроні, тім'я, лоб та наличник.
На боках голови, розміщені складні, багатофасеткові очі, які можуть мати округлу або ниркоподібну форми, а іноді є розділеними на дві окремі половини. У деяких родин жуків наявні ще й прості вічка на тім'ї.
Шиєю називається та частина головної капсули, яка входить в груди. Та частина голови, що безпосередньо примикає до шиї називається потилицею і має випуклу форму. Обабіч потилиці наявні характерні виступи — скроні.
Між очима, вище потилиці розміщене тім'я, яке на передній частині переходить у лоб. На лобі, між очима, розташовані вусики (антени) — це багаточленикові придатки, що виконують функції органів дотику й нюху, основи яких знаходяться у вусикових впадинах. Кількість члеників вусиків у різних родин твердокрилих дуже сильно варіює, так само як і їх форма. На передній частині лобу наявний шов, який відмежовує, від останнього, наличник. З боків голови, нижче очей, розміщені щоки.
До наличника причленяється верхня губа, яка частково прикриває мандибули і решту структур ротового апарата. Ротові придатки не занурені в ротову капсулу й складаються, окрім згаданої нечленистої непарної верхньої губи (лябрум), також із пари нечленистих верхніх щелеп (мандибули), пари нижніх щелеп (максили) та непарної нижньої губи (лябіум), яка утворилася в результаті злиття другої пари нижніх щелеп. Нижні щелепи та нижня губа двочленикові, несуть по парі органів дотику й смаку — щупиків. Для жуків характерний гризучий — найбільш примітивний, ротовий апарат.

Ротові органи жуків пристосовані для подрібнення твердої їжі — верхні щелепи мають жуйний край з розвиненими зубцями. Зубці лівої щелепи входять у заглибини правої. Максили складаються з основного членика та стовпчика, на якому є пара нижньощелепних щупиків; він закінчується рухомо причленованими жуйними лопатями — зовнішньою та внутрішньою. Нижня губа має основний членик, або підборіддя, та дистальний членик, на якому є дві пари жуйних лопатей. До ротового апарату належить також м'ясистий виріст ротової порожнини — гіпофаринкс. На нижній частині голови між шиєю та ротовим апаратом знаходиться горло. Його передня і найвужча частина, яка прилягає до нижньої губи називається субментумом або підборідком, а поздовжні боріздки біля його основи — горлові шви.
Будова грудей

Груди складаються з трьох сегментів, з яких найбільш розвиненим є перший. З дорзальної сторони вони називаються спинками: передньо-, середньо- та задньоспинкою. Кутикула кожного сегменту — це кільце, що поділяється на чотири склерити: спинний — терґіт, грудний — стерніт та два бічні — плейрити.
Передньоспинка часто має бокові шипи, вирости або горбики. Зверху помітною є лише передньоспинка, а середньоспинка, окрім щитка, та задньоспинка приховані надкрилами. Надкрила кріпляться до середньоспинки, а до задньоспинки — перетинчасті крила, призначені для польоту. Частина надкрил, яка заходить на нижню сторону тіла, називається епіплеврами. З нижньої сторони груди складаються з передньо-, середньо- та задньогрудей. Кожен з цих сегментів, у свою чергу, складається з центральної частини — диску, відмежованих від нього бічної — епістерн, та задньої — епімер, сторін. В задній частині кожного сегменту грудей знаходяться тазикові впадини, в яких розташовані, власне, тазики — частина кінцівки.
Будова кінцівок
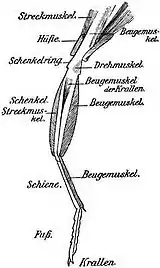

Кожний грудний сегмент має пару кінцівок. Кінцівки складаються з п'яти частин: тазиків, вертлюгів, стегон, гомілок та члеників лапок. Тазик — це короткий базальний членик, за допомогою якого нога рухомо прикріплюється до плейрита, вони можуть мати кулеподібну, конусоподібну або поперечно-поздовжню форму. Ця ознака дуже сильно варіює не лише у різних родин, а й у межах однієї родини. Тазик зі стегном сполучає вертлюг. Суглоби між тазиком і грудьми й тазиком і вертлюгом рухаються в різних площинах, забезпечуючи рухомість ноги. Стегно — найпотужніший і, як правило, найбільший членик кінцівки жука. До вершини стегна приєднується гомілка, а до неї лапка з члениками. На вершині гомілки розташовані шпори, зазвичай дві, а в деяких видів — по одній. Лапка складається, переважно, із 5-и члеників, хоча у різних родин цей критерій варіює, що є однією із систематичних ознак. Останній членик лапки, у всіх жуків, несе два кігтики. Залежно від способу життя в жуків розвинулися різні типи ніг: ходильні, копальні, стрибальні, плавальні тощо (докладніше див. статтю «Ноги комах»).
Будова крил
Крила — це бічні складки тіла, розташовані на середньо- та задньогрудях. Їх дві пари: передні та задні. У жуків передні крила перетворилися на тверді хітинізовані надкрила. Крило складається з двох стінок — верхньої та нижньої. Кожна стінка утворена шаром гіподерми, зовні вкритої більш-менш розвиненою кутикулою. Між стінками є вузенька щілина (частина міксоцелю — змішаної порожнини тіла), заповнена гемолімфою. Крило має систему хітинових трубочок-жилок. Жилки виконують опорну функцію; в них міститься гемолімфа, проходять трахеї та нерви до клітин крила.
Крила рухомо прикріплені до грудей між тергітом та плейритом за допомогою складної системи склеритів та мембран. Поблизу від місця прикріплення крило спирається на виріст плейрита — стовпчик, який є для нього точкою опори й утворює важіль з коротким осьовим і довгим кінцевим плечима.
Будова черевця
Черевце складається з 5-9-ти вільних кілець, які з нижньої сторони називаються терґітами, а з верхньої — прикритої крилами, стернітами. Останній членик черевця називається пігідієм.

Анатомія
Покриви тіла
Жуки мають три основні елементи покривів тіла: кутикулу, гіподерму і базальну мембрану. Кутикула утворює зовнішній скелет, що вкриває все тіло. Вона має поверхневий дуже тонкий шар — епікутикулу та внутрішній товстий — прокутикулу.
Епікутикула під світловий світловим мікроскопом має вигляд напівпрозорої поверхневої лінії, проте вона неоднорідна й складається із п'яти шарів, що різняться за хімічним складом. Найглибший шар епікутикули — протеїновий, над ним послідовно один на одному лежать шари твердого кутикуліну, восковий шар із гідрофобних ліпідних сполук і тонкий цементний. Останній утворює лакове покриття кутикули. Характерна особливість епікутикули — відсутність у її складі хітину.
_male_(14020453599).png.webp)
Прокутикула утворена двома шарами: м'якою безбарвною ендокутикулою, що прилягає до гіподермии, і твердою, забарвленою пігментами екзокутикулою. Ендокутикула містить полімерні молекули хітину, зв'язаного з білками. Вони лежать шарами, складеними з найтонших пластинок, причому їхній напрямок у кожному з шарів змінюється, що робить ендокутикулу еластичною й водночас міцною. Екзокутикула не має пластинчастої будови. Всю прокутикулу знизу доверху пронизують вертикальні порові канальці, всередині яких проходять тоненькі відростки гіподермальних клітин. Завдяки поровим канальцям відбуваються зв'язок гіподерми з кутикулою й утворення епікутикули під час линянь.
Гіподерма складається з одного шару призматичних клітин, між якими трапляються залозисті клітини, що утворюють одноклітинні або багатоклітинні шкірні залози, а також спеціалізовані клітини, які утворюють волоски та сенсили. Від порожнини тіла гіподерму відділяє базальна мембрана, яка не має клітинної будови.

У жуків розрізняють структурне й пігментне забарвлення. Структурне забарвлення пов'язане з деякими специфічними особливостями поверхневої структури кутикули — мікроскопічними реберцями, пластинками, лусочками тощо, які створюють ефекти інтерференції, дифракції та дисперсії світла. Це металево блискучі та переливчасті забарвлення деяких жуків, особливо у тропічних видів. Пігментне забарвлення зумовлене пігментами, які найчастіше містяться в екзокутикулі, рідше — в клітинах гіподерми чи жирового тіла. В багатьох жуків спостерігається поєднання структурного та пігментного забарвлення.
Найбільш поширеними пігментами жуків є меланіни — дають темно-коричневе, коричнево-червоне або чорне забарвлення. А також каротиноїди — жовте, жовтогаряче, червоне забарвлення; флавоноїди — жовтого, білого, червоного та коричневого кольорів.
Порожнина тіла

Порожнина тіла жуків, як і всіх комах, та й членистоногих загалом представлена змішаною порожниною тіла — міксоцелем. Під час ембріонального розвитку в них, як і в кільчастих червів, закладаються парні целомічні мішки, що мають метамерну будову. Пізніше стінки целомічних мішків руйнуються, розпадаючись на окремі клітини, а целомічні порожнини зливаються із залишками первинної порожнини тіла, утворюючи змішану порожнину тіла — міксоцель, що не має власної клітинної вистилки. Це система лакунарних або щілиноподібних порожнин між внутрішніми органами. Із мезодермальних клітин стінок целомічних мішків згодом утворюються мускулатура, клітини крові, жирове тіло та інші мезодермальні утвори. У міксоцелі циркулює рідина, що зветься гемолімфою. Вона є одночасно і порожнинною рідиною, і кров'ю.
Міксоцель — поділена двома поздовжніми горизонтальними перетинками — діафрагмами на три відділи (синуси). Верхня діафрагма відділяє верхній, або перикардіальний синус, в якому розташована спинна кровоносна судина. Нижня діафрагма відділяє нижній, або перинейральний синус, де міститься черевний нервовий ланцюжок. Середній синус лежить між діафрагмами; він називається вісцеральним; у ньому містяться травна, видільна й статева системи, а також більша частина жирового тіла. Порожнина тіла заповнена гемолімфою.
Центральна нервова система

Центральна нервова система жуків, як і інших комах, складається з парного надглоткового ганглія, або головного мозку, навкологлоткових конектив та черевного нервового ланцюжка. Перший ганглій ланцюжка — підглотковий — лежить разом із надглотковим у голові, решта — в тулубі.
Надглотковий ганглій складається з трьох злитих разом гангліїв: протоцеребрума, дейтоцеребрума та тритоцеребрума. Протоцеребрум розвинений краще, ніж інші, й має найскладнішу будову. У ньому розрізняють кілька гангліозних центрів, серед яких найкраще розвинена пара стебельчастих, або грибоподібних, тіл — вищого асоціативного та координуючого центру нервової системи. Крім того, в протоцеребрумі міститься пара великих зорових часток, які іннервують складні очі. Дейтоцеребрум — середній мозок — містить парні нюхові центри, він іннервує антени. Тритоцеребрум — задній мозок — іннервує верхню губу. З ним пов'язана вегетативна (симпатична) нервова система. Підглотковий ганглій іннервує ротові органи та слинні залози. Черевний нервовий ланцюжок злиттими грудними і черевними гангліїями.
Вегетативна нервова система
Крім центральної, у жуків добре розвинена вегетативна нервова система. Вона складається з трьох відділів: стомато-гастрального, вентрального, або черевного, та каудального. Стоматогастральний відділ складається з кількох самостійних гангліїв і нервів (фронтальний, потиличний, шлунковий ганглії, поворотний нерв), але має зв'язки з мозком. Стоматогастральна система іннервує серце й передню частину кишечника. Вентральний, або черевний, відділ складається з непарного нерва, який тягнеться паралельно до черевного нервового ланцюжка вздовж усього тіла; його вищим центром є тритоцеребрум. Вентральний нерв іннервує дихальця, трахеї, жирове тіло; разом із гангліями центральної нервової системи він посилає нерви до м'язів, справляючи на них регулювальний вплив. В останньому черевному ганглії непарний нерв розпадається на дві гілки, які іннервують задню кишку й статеві органи, цю його частину називають каудальним відділом.
Органи чуття
Жуки мають добре розвинені органи чуття: механорецептори (сприймають дотик, вібрацію та звукові хвилі); терморецептори (реагують на зміну температури); гігрорецептори (реагують на вологу); хеморецептори (сприймають хімічні стимули); фоторецептори або очі (сприймають світлові подразнення). Очі у жуків складні, або фасеткові, до складу яких входить велика кількість, інколи кілька тисяч, вічок, або оматидіїв, що щільно прилягають одне до одного. Кожен оматидій сприймає тільки одну точку предмета, що знаходиться перед ним, у результаті чого фасеткове око дає зображення, що складається з безлічі окремих точок, тобто мозаїчне. Є ще пропріорецептори, які сигналізують нервовій системі про положення, деформацію та зміщення окремих ділянок тіла.
Травна система

Травна система жуків складається з трьох відділів: передньої, середньої та задньої кишок. Стінки всіх відділів кишечника утворені одношаровим епітелієм, зовні вкритим поздовжніми та кільцевими м'язовими волокнами, скорочення яких забезпечує рух їжі в кишечнику. Епітеліальні клітини передньої та задньої кишок на вільній поверхні вкриті кутикулярним шаром — інтимою.
Передня кишка складається з ротової порожнини, глотки, стравохода, вола та м'язового шлунка. У ротову порожнину відкриваються слинні залози, пов'язані з ротовими кінцівками. У жуків наявні мандибулярні, максилярні та лабіальні (нижньогубні) залози. Слина змочує їжу та піддає її початковій дії ферментів, які розщеплюють поліцукри (крохмаль, глікоген).
Глотка й стравохід забезпечують проковтування їжі та її проходження до вола. Воло є місцем нагромадження їжі та початкового її перетравлювання під дією ферментів слини та травного соку, що потрапляє сюди із середньої кишки.
Жувальний шлунок має потужні м'язи; він вистелений усередині товстою кутикулою з гострими зубцями або товстими щетинками. У жувальному шлунку перетирається тверда їжа.
Середня кишка у вигляді прямої трубки відокремлена від передньої кардіальним клапаном — згорткою, що звисає в порожнину кишечника. Середня кишка є основним місцем травлення та всмоктування. Задня кишка відділена від середньої пілоричним клапаном і складається з тонкої, товстої та прямої (ректум) кишок. На межі між середньою та задньою кишками в кишечник відкриваються тоненькі трубочки — мальпігієві судини, які виконують видільну функцію. Задня кишка, як звичайно, не бере участі в травленні й не має ферментів. її функції пов'язані з формуванням екскрементів, водним обміном, виділенням та осморегуляцією.
Кровоносна система

Кровоносна система жуків, які і в більшості трахейнодишних (Tracheata) дуже редукована через, майже, повну втрату гемолімфою функції транспорту газів. Від неї залишається спинна судина, розташована в перикардіальному синусі й підвішена за допомогою сполучнотканинних тяжів до спинної стінки тіла. Задня її частина — серце, передня — аорта. Серце складається з ряду послідовних камер і розташоване в черевній тагмі. Кожна камера серця має пару бічних отворів — остій з клапанами. Через них гемолімфа потрапляє з перикардія всередину серця. Клапани перешкоджають її зворотному руху. Задній кінець серця замкнений, передній — подовжений у трубчасту аорту, яка відкривається в міксоцель поблизу голови. До верхньої діафрагми та нижньої сторони кожної камери прикріплюється пара крилоподібних м'язів.
Гемолімфа комах складається з рідкої міжклітинної речовини — плазми та клітин — гемоцитів, які або плавають у плазмі, або нерухомо осідають на поверхні внутрішніх органів. У в 1 мм3 гемолімфи міститься від 10 000 до 100 000 клітин, а їх загальний об'єм досягає 10 % об'єму гемолімфи. Плазма гемолімфи — це водний розчин неорганічних та органічних речовин. У ній є неорганічні йони та амінокислоти, які беруть участь у підтримці водно-сольового балансу й осморегуляції. Плазма гемолімфи містить також вуглеводи, органічні кислоти, гліцерин, ліпіди, пептиди, білки та пігменти.

Гемоцити — це клітини мезодермального походження. Усі вони безбарвні й мають ядра. Розрізняють кілька типів гемоцитів: одні з них можуть утворювати псевдоподії й здійснювати фагоцитоз, інші — нагромаджують поживні речовини і транспортують їх у тканини. У різних ділянках тіла, переважно в жировому тілі, є скупчення недиференційованих клітин, які перетворюються на гемоцити й потрапляють у плазму. Гемолімфа утворює рідке внутрішнє середовище організму. Вона виконує деякі важливі функції: 1. транспорт поживних речовин, гормонів та інших біологічно активних речовин, а також продуктів обміну до відповідних органів, тканин і клітин; 2. захист організму від інфекційних та інвазійних захворювань; 3. підтримка сталості хіміко-фізичних властивостей внутрішнього середовища організму; 4. механічна функція — внаслідок гідростатичного тиску гемолімфи змінюється форма органів з м'якою кутикулою — розправляються крила в імаго після виходу з лялечки, розкручується хоботок метеликів тощо. Гемолімфа має здатність до зсідання. При ушкодженні покривів вона витікає назовні й утворює згусток з гемоцитів та плазми, який закриває рану.

Неспецифічні імунні реакції забезпечують фагоцитуючі гемоцити, а також плазма, де є комплекс ферментів та антибіотиків із широким спектром дії на різні мікроорганізми, наприклад, фермент лізоцим руйнує оболонки бактеріальних клітин. Крім того, гемоцити утворюють капсули навколо багатоклітинних паразитів (нематоди, личинки їздців тощо), що призводить до загибелі останніх. Специфічний імунітет для жуків і комах не характерний, антитіла в гемолімфі не утворюються. Гемолімфа деяких жуків отруйна й використовується для захисту від ворогів.
Тісно пов'язане з гемолімфою жирове тіло, яке разом з нею утворює внутрішнє середовище організму. Ця пухка тканина мезодермального походження складається з численних лопатей між внутрішніми органами. Клітини жирового тіла за будовою та походженням близькі до гемоцитів. Більшість клітин жирового тіла становлять трофоцити. Основною функцією жирового тіла є нагромадження резервів поживних речовин на личинковій фазі розвитку та забезпечення ними організму на час метаморфозу, діапаузи, голодування, дозрівання статевих продуктів.
Дихальна система

Дихальна система жуків представлена трахеями. Трахеї — це розгалужені трубки ектодермального походження, складаються з одношарового епітелію й вистелені кутикулою. Трахеї відкриваються назовні кількома парами дихалець — стигм. Дві пари стигм розташовані відповідно на середньо- та задньогрудях, та перших п'яти сегментах черевця. Стигми мають замикальний апарат, який обслуговується двома м'язами й має спеціальну систему фільтрації повітря, побудовану з численних розгалужених щетинок. Кожне дихальце обслуговує три поперечні трахеї, з'єднані між собою трьома парами поздовжніх трахей від них відходять розгалуження до всіх органів. Вони закінчуються тоненькими трубочками діаметром 1-2 мкм — трахеолами. Кінці трахеол або лежать на поверхні окремих клітин, або входять всередину. Кисень із трахеол безпосередньо дифундує в клітини, а вуглекислий газ — із тканин у трахеоли.
Видільна система
Видільна система представлена мальпігієвими судинами, задньою кишкою, уратними клітинами жирового тіла, перикардіальними клітинами та ін. Основними видільними органами є мальпігієві судини й задня кишка, що функціонують як єдине ціле. Це довгі тоненькі трубочки, які впадають у кишечник на межі середньої та задньої кишок. Протилежні сліпозамкнені кінці їх вільно плавають у гемолімфі. Стінки судин утворені одношаровим епітелієм, зовні вкритим базальною мембраною та м'язовими волокнами. Скорочення м'язів спричиняють рух судин у гемолімфі, мальпігієві судини всмоктують гемолімфу з продуктами обміну. З уратів — солі сечової кислоти — у мальпігієвих судинах утворюється малорозчинна сечова кислота. У задній кишці ректальні сосочки вилучають із цієї рідини й повертають до гемолімфи більшу частину води, поживні речовини та іони неорганічних сполук. Зневоднені кристали сечової кислоти разом із фекаліями виводяться назовні через анальний отвір.
У жировому тілі, крім клітин-трофоцитів, які запасають поживні речовини, трапляються також уратні клітини, що нагромаджують сечову кислоту. Нагромаджувальна функція цих клітин має важливе значення в ті періоди розвитку комах, коли виведення екскретів назовні неможливе. Перикардіальні клітини, що оточують спинну кровоносну судину, здатні поглинати великі білкові молекули та різні колоїдні частинки, які потрапляють у гемолімфу.
Статева система

Статева система самиці
Статева система самиці складається з пари яєчників, пари яйцепроводів, непарного яйцепроводу, придаткових статевих залоз, сім'яприймача. Кожен яєчник складається з яйцевих трубок, які у свою чергу складаються з верхівкової частини — гермарія та базальної — вітелярія. У гермарії утворюються й розмножуються первинні статеві клітини, з яких потім виходять ооцити й живильні клітини. Сформовані ооцити потрапляють у вітелярій, де вони дозрівають, нагромаджують поживні речовини у вигляді жовтка й стають яйцеклітинами. Вітелярій поділений на ряд яйцевих камер. У кожній із них є лише одна яйцеклітина. В міру росту вона наближається до виходу з вітелярія, тому останні яйцеві камери найбільші; у них містяться вже готові до відкладання яйця. Ріст і розвиток яйцеклітини відбуваються за рахунок надходження до неї поживних речовин, що виробляються в живильних клітинах, а також у фолікулярному епітелії, який формує стінки яйцевих камер. Після закінчення розвитку яйця фолікулярний епітелій виділяє захисну оболонку — хоріон.
Дозрілі яйця з яйцевих трубок потрапляють у парні яйцепроводи, потім — у непарний яйцепровід, а звідти через статевий отвір виходять назовні. У непарний яйцепровід впадає протока сім'яприймача, призначеного для зберігання сперматозоїдів, які потрапляють до нього під час парування. Яйце запліднюється, проходячи через непарний яйцепровід під час відкладання яєць; сперматозоїди в цей час виходять із сім'яприймача й проникають у яйце.
Статева система самця
Статева система самця складається з пари сім'яників, пари сім'япроводів, сім'явипорскувального каналу, придаткових статевих залоз і копулятивного органа. Сперматозоїди з сім'яників потрапляють до сім'япроводів, з них — у сім'явипорскувальний канал, і при паруванні виводяться назовні. Часто копулятивний орган утворює зовнішні статеві придатки — геніталії. Придаткові залози, які відкриваються у сім'явипорскувальний канал.
Онтогенез
Ембріогенез жуків

Ембріональний розвиток жуків має свої особливості. Яйцеклітини у них багаті на жовток, який займає всю центральну частину яйця. Тому дробіння в них часткове, поверхневе; у результаті утворюється поверхневий шар однакових клітин — бластодерма, яка покриває центральну масу жовтка. На майбутній черевній стороні зародка клітини бластодерми починають інтенсивніше ділитися, бластодерма потовщується. Ця ділянка зветься зародковою смужкою. Пізніше за її рахунок формується більша частина зародка.
У зародковій смужці відбувається гаструляція (найчастіше інвагінацією або епіболією), причому спочатку утворюються лише два зародкових листки: ектодерма та мезодерма. Ектодерма дає початок зовнішнім покривам тіла, трахеям, нервовій системі, статевим протокам, передньому та задньому відділам кишечника, що виникають як вгинання покривів на передньому та задньому кінцях зародка.
Із мезодерми утворюється мускулатура, жирове тіло, серце з аортою, гемоцити. Мезодерма бере участь також у розвитку статевої системи. Статевий зачаток, як правило, відокремлюється значно раніше від інших тканин у вигляді парної групи клітин біля заднього кінця зародка. Ентодерма, з якої походить середня кишка, утворюється за рахунок невеличких зачатків на дні передньої та задньої кишок, які розростаються назустріч один одному, доки не утвориться суцільна трубка — кишечник. Паралельно з розростанням зародка в оболонках починається його сегментація. На головному кінці з'являються зачатки очей, відокремлюються сегменти голови, грудей та черевця. З'являються зачатки кінцівок, у тому числі й на черевці. Пізніше черевні кінцівки зникають. Мезодерма при цьому теж сегментується на кілька целомічних мішечків, які потім розпадаються.
Урешті-решт увесь жовток вичерпується, і сформований зародок заповнює все яйце. Він прогризає чи прориває оболонку яйця й виходить назовні. Постембріональний розвиток комах відбувається з метаморфозом і має епіморфний характер. Це означає, що з яйця виходить личинка з усіма сегментами, яка відрізняється від дорослої особини — імаго.
Личинка
.jpg.webp)

.jpg.webp)
Для жуків властива голометаболія, при якому комаха проходить фази яйця, личинки, лялечки, імаго. З яйця виходить личинка, яка різко відрізняється від імаго більшою гомономністю метамерії, відсутністю зовнішніх зачатків крил, слабшим розвитком органів чуття. Закінчивши свій ріст, личинка останньої стадії припиняє живлення, стає нерухомою, линяє востаннє й перетворюється на лялечку.
Лялечка
У жуків лялечка належить до, так званих, покритих лялечок, у яких імагінальні придатки тісно зрощені з тілом унаслідок того, що личинка при останньому линянні виділяє секрет, який при затвердінні вкриває лялечку твердою оболонкою. Лялечка нерухома, але це тільки зовнішнє враження: на цій фазі відбуваються інтенсивні процеси внутрішньої перебудови — гістоліз та гістогенез. Оскільки характер руху імаго, особливо політ, і його живлення інакші, ніж у личинкової фази, то і м'язи, і травна система, кінцівки та інші личинкові органи мають зовсім перебудуватися. Більшість органів личинки руйнується; цей процес зветься гістолізом. Він відбувається фагоцитозом, автолізом (самоперетравлення), а також за допомогою ферментів. Гемоцити виділяють у гемолімфу ферменти, які руйнують тканини; їх залишки фагоцитуються гемоцитами. Крім того, в деяких тканинах відбувається автоліз.
Коли гістоліз досягає свого піку, внутрішні органи лялечки перетворюються на напіврідку масу, яка складається з гемолімфи, збагаченої продуктами розпаду. Не руйнуються лише нервова, статева системи та спинна кровоносна судина. Нервова система може доповнюватися новими клітинами, в ній можуть концентруватися ганглії, проте вона ніколи не втрачає своєї цілісності.
Паралельно з гістолізом відбувається гістогенез — побудова імагінальних органів. Органи дорослої комахи формуються з особливих зачатків — імаґінальних дисків. Це невеличкі скупчення недиференційованих клітин, розташовані в певних місцях тіла личинки. Кожен імагінальний диск має своє призначення: є диски, з яких утворюються крила, кінцівки, певні ділянки кишечника, фасеткові очі тощо. Імагінальні диски закладаються ще під час ембріонального розвитку або в ранніх личинок; протягом личинкового розвитку вони ростуть, але не диференціюються. Тільки в личинки останньої стадії перед залялькуванням їхні клітини диференціюються; в лялечки вони розгортаються, утворюючи імагінальні органи. Важливу роль у метаморфозі відіграє жирове тіло. На личинковій фазі в ньому нагромаджуються поживні речовини (жири, білки, вуглеводи), що використовуються як пластичний матеріал і джерело енергії при побудові імагінальних органів.
Метаморфоз комах відбувається під контролем гормональної (ендокринної) системи. Як уже зазначалося, метаморфоз регулює нейроендокринний комплекс органів, до якого входять нейросекреторні клітини мозку, кардіальні тіла, прилеглі тіла та парна передньогрудна, або проторакальна, залоза. Нейросекреторні клітини мозку виробляють активаційний гормон. Через довгі аксони цих клітин він надходить до кардіальних тіл, а з них — у гемолімфу. У личинок активаційний гормон стимулює роботу проторакальних залоз, які починають виділяти линяльний гормон — екдізон. Останній впливає на клітини гіподерми, спричиняючи спочатку синтез ферментів, які розчиняють стару кутикулу, а пізніше — синтез матеріалу для побудови нової кутикули. Крім того, екдізон стимулює ріст і диференціацію всіх тканин личинки, особливо в гонадах, імагінальних дисках тощо.
Паралельно з екдізоном при личинкових линяннях виділяється ювенільний (юнацький) гормон, що гальмує метаморфоз, тобто перешкоджає линянню личинки на дорослу фазу. Наявність ювенільного гормону в гемолімфі визначає характер линяння. При високій концентрації ювенільного гормону екдізон гальмує розвиток імагінальних органів та стимулює секрецію личинкової кутикули, і линяння завершується появою чергової личинкової стадії. Зниження концентрації ювенільного гормону визначає линяння личинки на лялечку. Припинення надходження ювенільного гормону в гемолімфу обумовлює линяння лялечки на імаго.
Філогенія

Найдавніші палеонтологічні знахідки жуків відносяться до кінця кам'яновугільного періоду палеозойської ери — 320—300 млн років тому. Із сучасних 166 родин жуків, 67 відомі з Палеозойської та Мезозойської ер, і лише 19 із них є такими, що вимерли до початку Кайнозойської ери.
Вважається, що вихідною групою, з якої у подальшому сформувався ряд Твердокрилих, була родина Щекардоколіїїди (Tschekardocoleidae), котрих відносять до Першожуків (Protocoleoptera). Протоколеоптери або Першожуки виникли у кам'яновугільному періоді Палеозою. До початку Пермського періоду від них виокремилась еволюційна гілка Жуків-Пермокупедоідей (Permocupedoidea), яких виокремлюють у ранг надродини. Пермокупедоідеї проіснували до початку Крейдового періоду (120—145 млн років тому) Мезозойської ери і вимерли. Проте, ще у нижньому Тріасовому періоді — 240—250 млн років тому, від цієї групи відокремилась еволюційно дуже успішна філа Жуків-Архостемат (Archostemata), які живуть і нині. Архостемати утворюють окремий підряд у ряді Жуків і є найдавнішими представниками цього ряду. Для Архостемат характерний розвиток личинок у деревині, яка виступає для них джерелом їжі, а оскільки умови середовища деревини в ході еволюції змінювались мало, то жуки цього підряду не зазнавали впливу різких змін клімату і не вимерли. Раннім Тріасом датуються також і перші знахідки ушкоджень деревини комахами — у скам'янілих стовбурах дерев чітко видні ходи личинок.
Пермокупедоідеї включали родину Жуків-Адемосинід (Ademosynidae), яка вимерла разом із ними в Крейдовому періоді, проте, у середньому Тріасі ця родина дала початок, нині найоб'ємнішому за кількістю видів, підряду Всеїдних жуків (Polyphaga).
До пермських жуків також відносяться ще дві надродини: Ромбоколеоідей (Rhombocoleoidea) та Азіоколеоідей (Asiocoleoidea), якщо остання надродина проіснувала до початку Юрського періоду (170—190 млн років тому) і вимерла, то перша у верхньому Тріасі розвинулась у інфраряд Шізофоріформ (Schizophoriformia), від якого походять два важливі підряди жуків — Змішаноїдні (Myxophaga) та Хижі (Adephaga), що успішно існують й сьогодні.
Походження

Палеонтологічна наука, на нинішній день, не може дати якоїсь достовірної відповіді на питання про походження жуків, тому воно залишається достеменно невідомим. Перехідних форм між ними і якимись іншими групами комах, що мають повне перетворення, наразі, не виявлено, хоча існує певна схожість між найдревнішими жуками та комахами із неповним перетворенням. У геологічних відкладах кам'яновугільного та пермського періодів пелеозою зустрічаються дві групи комах дуже схожих на древніх жуків — це Протелітроптери й Таргани, що викликає дуже багато плутанини в ідентифікації древніх жуків.
Знахідки жуків, що датуються як найдревніші, вказуються з відкладів асельського ярусу Пермського періоду знаходженням у Нідермошелі, що поблизу Майнцу, Німеччина. Проте, багато вчених сумнівається у їх правдивості. Науковозадокументованими, найдревнішими, є знахідки жуків сакмарського ярусу Пермського періоду із Босковицького Грабена, Східна Чехія. ці знахідки є дуже різноманітними і описані у 8-и родах, приналежних до родини Щекардоколеїди (Tshekardocoleidae). Їхні надкрила характеризуються ґратчастою структурою із жилкуванням схожим на жилкування древніх Мегалоптер, з якими, можливо, жуки мають спільне походження. Крила також не здатні були складатись на вершині, на відміну від сучасних жуків, а лежали вздовж черевця і надкрила були значно довшими за черевце. Інші ознаки, як наприклад, рухомі передньогруди, злиті і приховані стерніти задньогрудей, занурені в груди тазики задніх ніг, склеротизовані базальні членики черевця тощо, не відрізнялись від сучасних жуків.
Знахідки жуків з кунґурського ярусу Пермського періоду на Уралі, біля Черкади в Пермській області, Росія, приналежних до тієї ж родини Щекардоколеїд, свідчать про те, що структура надкрила значно змінюється — жилки випрямлені з меншою кількістю комірок. У середині Пермського періоду надкрила жуків структурно не відрізняються від сучасних — жилки розширюються, цілком заповнюючи простір комірок, а останні зменшуються до порожнистих стовпчиків-колумел. Процес становлення надкрила відбувається двома шляхами. Перший — у Жуків-Купеїдів жилки переднадкрил утворюють механічно досконалу ґратчасту структуру, і тільки після цього починають розростатись, витісняючи комірки, які набувають вигляду штрихових боріздок. Другий — у Жуків-Схізофороїдів розростання жилок відбувається без проміжних стадій спеціалізованого жилкування. Одночасно у всіх групах жуків виникають гладенькі надкрила — вважається, що то є пристосуванням до їх життя у водному середовищі, адже гладке надкрило спричинює меншу турбулентність водного потоку і забезпечує ефективний рух у товщі води.
На початку Пермського періоду жуки є надзвичайно рідкісними тваринами, навіть у масових захороненнях комах, вони складають не більше 0,1-0,6 % від всієї ентомофауни. А у численних відкладах артинського ярусу Пермського періоду Північної Америки, де віднайдено багато залишків комах, жуки взагалі відсутні, і це тоді, як Європа і Північна Америка складали єдиний материк.
Екологія
Твердокрилі, як одна із найдревніших та найуспішніших груп класу комахи (Insecta), зуміли заселити всі можливі суходільні та водні екологічні ніші. Вони розповсюджені на всій Землі: від найбідніших пустель до дощових тропічних лісів, від екваторіальної до полярних зон; немає жуків лише у місцях багаторічних льодовиків, таких як арктичні, антарктичні і високогірні. В гірських місцевостях розповсюджені до субнівального та нівального поясів. У Карпатах їх можна зустріти від передгір'їв до найвищих вершин; на Кавказі жуки трапляються до висот понад 3000 м; в горах Середньої Азії — 4500, а у Гімалаях та тропічних гірських масивах — до 5000-5500 м.
Різноманітність екологічних груп жуків є надзвичайно великою, а ніші, які вони заселяють, — надзвичайно різноманітні. Велика група видів жуків заселяє верхні шари ґрунту і називається ґрунтовою мезофауною (Туруни, Жуки-хижаки, Слоники та ін.), причому личинки і лялечки там проводять увесь свій час, а імаго, хоча б на короткий період, виходить на поверхню. Ще більшою групою жуків є види, які заселяють рослини. Багато видів живе на листках і стеблах рослин, якими вони живляться (Листоїди, Ковалики, Слоники тощо), інші заселяють деревину дерев (Вусачі, Златки й ін.) та простір під корою (Плоскотілки, Короїди, Лубоїди і т. д.), ще інші поселяються в коренях і на їх поверхні (Пластинчастовусі, деякі Слоники й Вусачі), на квітах та в плодах (Слоники, Насіннєїди тощо). Велика кількість видів жуків мешкає в прибережних біотопах прісних водойм. Варто виокремити групу прісноводних жуків-гідробіонтів (Вертячки, Плавунці, Водолюби та ін.), які ведуть, переважно, хижий спосіб життя. Велика група хижих жуків заселяє травостій та лісову підстилку (Туруни, Жуки-хижаки), інші хижаки полюють на листі рослин (Сонечка, М'якотілки та ін.). Значна, за обсягом видів, група жуків мешкає в продуктах життєдіяльності тварин (Гнойовики, Пластинчастовусі, Карапузики тощо) та їх трупах (Мертвоїди й ін.)
Екологічні групи жуків за типами живлення імаго та личинок:
- Гідробіонтні хижаки — види, що живляться тваринною їжею, полюючи у водних екосистемах;
- Педобіонтні хижаки — види, що живляться тваринною їжею, полюючи на поверхні ґрунту;
- Фітобіонтні хижаки — види, що живляться тваринною їжею, полюючи на поверхні рослин;
- Листо- та хвоєгризні фітофаги — види, що живляться рослинною їжею, об'їдаючи листки або хвою;
- Ксилофаги — види, що живляться рослинною їжею, поїдаючи деревину;
- Ризофаги — види, що живляться рослинною їжею, обгризаючи корені або живуть в їх середині;
- Антофаги — види, що живляться рослинною їжею, поїдаючи квітки або їх частини;
- Карпофаги — види, що живляться рослинною їжею, поселяючись всередині плодів та насіння;
- Міцетофаги — види, що живляться плодовими тілами або міцелієм грибів;
- Детритофаги — види, що живляться відмерлими рослинними рештками;
- Некрофаги — види, що живляться рештками мертвих тварин;
- Копрофаги — види, що живляться фекаліями тварин.
- Паразитоїди — види, личинки яких живляться частинами личинок або лялечок інших комах, на їх поверхні;
- Паразити — види, личинки яких живляться в середині личинок або лялечок інших комах
Екологічні групи жуків за типами біотопів:
- Гідробіонти — види, що заселяють водойми;
- Педобіонти — види, що заселяють ґрунт;
- Галлобіонти — види, що заселяють дуже засолений ґрунт;
- Галлофіли — види, що віддають перевагу засоленим ґрунтам;
- Фітобіонти — види, що заселяють рослини;
- Ксилобіонти — види, що заселяють деревину;
- Мірмекофіли — види, що заселяють мурашники;
- Термітофіли — види, що заселяють термітники;
- Нідіколи — види, що заселяють нори ссавців або гнізда птахів;
- Троглобіонти — види, що заселяють печери;
- Троглофіли — види, що тимчасово заселяють печери та порожнини під великими каменями;
- Синантропи — види, що заселяють людські оселі і будівлі;
Систематика
Колеоптерологічні колекції

Жуки є найбільш популярною, звичайно, після метеликів, групою комах для приватних колекціонерів та науковців. Це зумовлено, в першу чергу, величезною кількістю видів, їх розповсюдженістю на всій Землі, а також великою кількістю представлених серед них форм, розмірів та кольорів. Саме тому колекціонування жуків набуло значного розповсюдження. Колекції варто розмежувати на приватні та наукові. Приватні колеоптерологічні колекції, часто носять аматорський характер і орієнтуються на естетичність та ефектність зібраних екземплярів. Жуків до таких колекцій підбирають за розмірами — чим більші, тим краще — та кольоровою гаммою. Найбільш популярними об'єктами колекціонування є Жуки-туруни із родів Турун, Красотіл та ін., які характеризуються значними — 3-5 см — розмірами тіла і широкою гаммою кольорів. Окрім них найчастіше ще колекціонують Жуків-вусачів та Жуків-скарабеїдів. З часом, за звичай, інтереси приватних колекціонерів розповсюджуються й на дрібніших представників з інших родин. Іноді, такі колекції переростають у солідні наукові фонди.
Наукові колеоптерологічні колекції — це фонди музеїв, університетів, науково-дослідних інститутів, карантинних служб та інших наукових установ. Такі колекції збираються науковцями, які проводять спеціалізовані фауністичні, еволюційні та екологічні дослідження жуків, і, як правило, є національним науковим та культурним надбанням з відповідно оформленою документацією та регламентованими правилами користування. Колекції такого типу поділяються на виставкові — призначені для всезагального огляду, та фондові, доступ до яких мають лише наукові співробітники. У фондових колекціях, найчастіше зберігається серійний типовий матеріал, за яким здійснено першоопис певного виду жуків, і всі подальші знахідки цього виду, для точності визначення, порівнюються з типовими: голотипом чи паратипом.
Посилання
- Велика історія жуків(укр.)
Примітки
- А. Ф. Бартенев, А. Г. Шатровский, Д. В. Вовк. Обзор семейств жуков (Coleoptera) Украины. Часть 2. Polyphaga (Staphyliniformia: Hydrophiloidea; Scarabaeiformia: Scarabaeoidea) // Известия Харьковского энтомологического общества. - 1997. - Т. 5, Вып. 1. - С. 5–21. -
Література
- Гурьева Е. Л., Крыжановский О. Л. (ред.). 1965 Определитель насекомых европейской части СССР. Том 2. Жесткокрылые и веерокрылые. М.-Л., Наука. 668 с.
- Андрій М. Заморока Курс лекцій із зоології — Івано-Франківськ, 2006.
- Пономаренко А. Г. Историческое развитие жесткокрылых насекомых. Автореф. дисс. докт. биол. наук — М. — 1983 — 47 сс.
- Crowson R.A. 1981 The Biology of the Coleoptera. London, Academic Press. 802 p.
- Kirejtshuk A.G. Evolution of mode of life as the basis for division of the beetles into groups of high taxonomic rank // In: Zunino M., Belles X. & Blas M. (eds.) Advances in Coleopterology. — Barcelona: Assoc. Europ. Coleopt. — 1992/1991 — P. 249—261.
- Lawrence J.F., Newton A.F. 1995 Families and subfamilies of Coleoptera (with selected genera, notes, references and data on family-group names) // In: Biology, Phylogeny, and Classification of Coleoptera. Eds. J. Pakaluk and S.A. Slipinski. Warszawa, 1995: 779—1006.
- Ponomarenko A.G. The geological history of beetles // in: J. Pakaluk & S.A. S'lipin'ski (eds.). Biology, Phylogeny, and Classification of Coleoptera. Papers celebrating the 80th Birthday of Roy A. Crowson. — Warszawa: Muz. Inst. Zool. PAN — 1995 — Vol. 1 — P. 155—172.
|
|
Ця стаття належить до добрих статей української Вікіпедії. |