Аденозиндезамінази РНК
Аденозин-дезамінази РНК, ADAR — родина білків-ферментів дезаміназ, які відщеплюють аміногрупу від азотистої основи аденіну, яка знаходиться у полінуклеотидному ланцюгу РНК. Аденозин-дезамінази розпізнають дволанцюгову ділянку РНК і є основними ферментами, які забезпечують процес редагування РНК у хребетних, зокрема у ссавців і людини.
| Аденозин-дезаміназа РНК 1 | |
|---|---|
 ADAR1 людини | |
| Ідентифікатори | |
| Символ | ADAR |
| Інші символи | ADAR1; AGS6; DRADA; DSH; DSRAD; G1P1; IFI-4; IFI4; K88DSRBP; P136 |
| Entrez | |
| OMIM | |
| RefSeq | |
| UniProt | |
| Інша інформація | |
| Шифр КФ | |
| Локус | Хр. 1 |
| Аденозин-дезаміназа РНК 2 | |
|---|---|
 ADARB1 людини | |
| Ідентифікатори | |
| Символ | ADARB1 |
| Інші символи | RED2; ADAR3 |
| Entrez | |
| OMIM | |
| RefSeq | |
| UniProt | |
| Інша інформація | |
| Шифр КФ | |
| Локус | Хр. 21 |
| Аденозин-дезаміназа РНК 3 | |
|---|---|
| Ідентифікатори | |
| Символ | ADARB2 |
| Entrez | |
| OMIM | |
| RefSeq | |
| Інша інформація | |
| Локус | Хр. 10 |
За амінокислотною послідовністю й рентгенограмами білкових кристалів вони принципово відрізняються від інших аденозиндезаміназ (ADA), що відщеплюють аміногрупу від аденозинмонофосфату (АМФ). Деяку гомологію спостерігають між генами ADAR та цитозиндезаміназ, що може свідчити про їх походження від спільного гену-предка[1]. Утворений внаслідок дезамінування нуклеотид інозин розпізнається системами трансляції, РНК-інтерференції, зворотної транскрипції та іншими молекулярними машинами, побудованими на принципі комплементарності, як гуанозин.
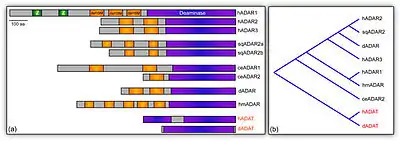
До родини аденозин-дезаміназ у ссавців належать 3 гени: ADAR1, ADAR2, ADAR3. Білок ADAR3 присутній лише в головному мозку, тоді як ADAR1 і ADAR2 наявні в усіх тканинах, хоча більше їх також у ЦНС.
Структура
На N-кінці білка ADAR1 знаходяться 2 РНК-зв'язувальних домени, а також 2 Z-ДНК-зв'язувальні сайти. C-кінець містить каталітичний дезаміназний домен, який у ADARB2 є неактивним.[2]
Аденозин-дезамінази діють у формі димерів. Кожен із білків ADAR має у своєму складі РНК-зв'язувальний домен, який містить іон цинку. Також в активному центрі ферменту є молекула інозитол-гексафосфату (IP6). Білок ADAR1 людини має в своєму складі додатковий сайт зв'язування з іоном цинку, який сприяє реакції дезамінування аденозину[3].
Білок ADAR1 існує у вигляді декількох ізоформ. Ізоформа ADAR1p110 коротша, не містить ДНК-зв'язувального домену, представлена в багатьох тканинах та не потребує додаткових сигналів для транскрипції і трансляції (конститутивна форма). Натомість ADAR1p150 є повнорозмірним білком, який експресується під дією інтерферонів. У мишей ADAR1p150 присутній у всіх тканинах окрім головного мозку. ADAR1p150 переважно локалізований у цитоплазмі клітин, тоді як ADAR1p110 — у ядрі.[4]
Регуляція
Білки ADAR є мішенями фосфорилювання ферментами родини AKT-кіназ[5].
Функції
Редагування РНК
Аденозин-дезамінази взаємодіють із первинними транскриптами, найчастіше — до сплайсингу РНК. Мішенню є дволанцюгова РНК, зазвичай — спарені нуклеотиди у шпильках. Невідомо, чи існують послідовності, що є сприятливими до редагування, але є відомості щодо специфічної третинної й четвертинної укладки молекули РНК, яку використовують аденозин-дезамінази. Редагування відбувається в пре-мРНК, первинних транскриптах мікроРНК, у транскриптах повторів (наприклад, Alu-елементів). Редагування призводить до замін амінокислот у білку, що синтезується з мРНК, до створення або ліквідації сайтів для сплайсингу чи до зміни активності мікроРНК.
Вкорочений сплайс-варіант ферменту ADAR1 і білок ADAR2 містяться в ядрі та експресуються постійно. Білок ADAR1 із повною послідовністю (p150, масою 150 кілодальтон) рухається з ядра до цитоплазми й назад. Для білку ADAR3 людини не доведено дезаміназної активності[6].
Редагування А на І може відбуватися для всіх транскриптів (Q/R сайт глутаматних рецепторів — понад 99,9 % транскриптів редагуються) або лише для частини з них, тоді як інші транскрипти залишаються нередагованими. Регуляція редагування здійснюється багатьма факторами: інтерфероном, транскрипційними кофакторами, збиранням субодиниць білка в димери тощо. ADAR2 саморегулюється через негативний зворотний зв'язок: надлишок білка починає редагувати пре-мРНК самого ADAR2, викликаючи появу додаткового сайту сплайсингу й утворення вкороченого нефункціонального ферменту.
Досліди з нокауту генів аденозин-дезаміназ довели, що вони є життєво важливими для ссавців. Миші з вимкненим ADAR1 гинули ще в ембріональний період розвитку через масову втрату нейронів апоптозом і порушення системи кровотворення. Миші, позбавлені ADAR2, помирали одразу після народження через епілептичну активність мозку. Проте заміна лише одного нуклеотиду А на Г у сайті гену GluR2, що кодує глутамін, надавала змогу нокаутним мишам жити без таких порушень[6].
Частина сайтів редагування в мишей редагується переважно ADAR2, частина ADAR1, а інші редагуються обома ферментами[7].
У нематоди Caenorhabditis elegans білки ADAR задіяні в пригніченні активності ретротранспозонів та ендогенних вірусів[8].
РНК, що містить багато відредагованих інозинів, розпізнається білком SND1, який входить до складу РНК-індукованого комплексу заглушення біосинтезу білка. Це призводить до деградації такої РНК за допомогою ендонуклеази ENDOV.[4]
У дрозофіли попри наявність лише одного гену ADAR, гомологу ADAR2, відбувається активне редагування РНК[9].
Інші функції
ADAR1 без зв'язування з РНК взаємодіє з білком UPF1, який входить до складу сплайсосоми, та є необхідним для руйнування інтронів після сплайсінгу. ADAR1 регулює транскрипцію в B-лімфоцитах через взаємодію з білком ELAVL1.[4] Також ADAR1 зв'язується з протеїнкіназою PKR, пригнічуючи її противірусну дію. У зв'язуванні з PKR задіяні Z-ДНК–зв'язувальний домен та домен зв'язування для дволанцюгової РНК[10].
Походження та еволюція
Білки ADAR мають високий ступінь гомології з аденозин-дезаміназами тРНК (ADAT), а ті, у свою чергу, близькі до іншої родини ферментів — цитидиндезаміназ. При цьому не спостерігається жодної подібності у ADAR і аденозиндезаміназ пуринового обміну.
Гени ADAR наявні у більшості багатоклітинних тварин, хоча окремі групи втратили один чи обидва гени. Зокрема дрозофіли та інші комахи мають лише гомолог ADAR2.
Найбільшу кількість генів ADAR станом на 2018 рік мають реброплави, у геномі яких наявні 4 ізоформи аденозиндезаміназ. Ці ферменти є найбільш активними під час ембріогенезу. ADAR1 експресується на найвищому рівні під час раннього дроблення, тоді як ADAR2 і ADAR3 працюють під час формування війчастих гребінців.[11]
Очисний добір у більшості видів тварин прибирає ідеально комплементарні дволанцюгові РНК, а ті, що залишаються, чи нові підлягають потужному редагуванню за допомогою ADAR1[12].
Захворювання
Мутації
Мутації гену ADAR призводять до виникнення рідкісного аутосомно-домінантного спадкового захворювання — симетричного спадкового дисхроматозу, поширеного переважно серед японців, але відомого з європейської, індійської та карибської популяції.[6]
Також мутації в цьому гені можуть призводити до рідкісного автоімунного синдрому Екарді-Гут'єр[13].
Нейрологічні захворювання
У пацієнтів з хворобою Альцгеймера виявлено знижений рівень редагування аденозинів у РНК білків, пов'язаних з роботою головного мозку[14]. Більша варіабельність редагування спостерігалася в гіпокампі, зокрема в Q/R сайті[15].
Редагування РНК серотонінового рецептора HTR2C посилюється під дією інтерферона, оскільки в 4 сайтах забезпечується білком ADAR1[16] Зміни рівнів редагування пре-мРНК цього рецептора спостерігаються при шизофренії, бічному аміотрофічному склерозі, під дією хронічного стресу, викликаного соціальною поразкою.
Інфекційні захворювання
Оскільки білки ADAR здатні зв'язуватися й редагувати довгі дволанцюгові РНК, притаманні багатьом вірусам на різних стадіях розвитку, то їхньою функцією вважалася противірусна дія шляхом редагування й пошкодження вірусного геному чи транскриптому. Перші свідчення такої дії були показані на клітинних лініях для вірусу кору[17]. Однак упродовж 2010-х років накопичилося чимало досліджень, які виявили сприяння розмноженню вірусів з боку аденозиндезаміназ РНК. Зокрема це показано для вірусів кору, везикулярного стоматиту, ВІЛ, гепатиту D, Зіка.[10][18]
Білки ADAR можуть відігравати роль у мутуванні РНК-вірусів[19]. Також редагування РНК ВІЛ за допомогою ADAR1 не викликає суттєвих пошкоджень вірусних часточок та їхньої здатності інфікувати клітини[20].
Використання
Створено декілька штучних генетичних систем, елементом яких є аденозиндезамінази РНК, які здатні вибірково замінювати аденозин на інозин у РНК[21][22].
Див. також
Примітки
- Willemijn M. Gommans, Dylan E. Dupuis, Jill E. McCane, Nicholas E. Tatalias, Stefan Maas (2008). Diversifying Exon Code through A-to-I RNA Editing. У Smith, H. DNA RNA Editing. Wiley & Sons, Inc. с. 3–30.
- Savva, Yiannis A; Rieder, Leila E; Reenan, Robert A (2012). The ADAR protein family. Genome Biology 13 (12): 252. ISSN 1465-6906. doi:10.1186/gb-2012-13-12-252.
- Park, SeHee; Doherty, Erin E.; Xie, Yixuan; Padyana, Anil K.; Fang, Fang; Zhang, Yue; Karki, Agya; Lebrilla, Carlito B.; Siegel, Justin B.; Beal, Peter A. (2020). High-throughput mutagenesis reveals unique structural features of human ADAR1. Nature Communications 11 (1). ISSN 2041-1723. doi:10.1038/s41467-020-18862-2.
- Sinigaglia, Ketty; Wiatrek, Dagmara; Khan, Anzer; Michalik, David; Sambrani, Nagraj; Sedmík, Jiří; Vukić, Dragana; O'Connell, Mary A. та ін. (2019). ADAR RNA editing in innate immune response phasing, in circadian clocks and in sleep. Biochimica et Biophysica Acta (BBA) - Gene Regulatory Mechanisms 1862 (3): 356–369. ISSN 18749399. doi:10.1016/j.bbagrm.2018.10.011.
- Piazzi, Manuela; Bavelloni, Alberto; Gallo, Angela; Blalock, William L. (2020). AKT-Dependent Phosphorylation of ADAR1p110 and ADAR2 Represents a New and Important Link Between Cell Signaling and RNA Editing. DNA and Cell Biology 39 (3): 343–348. ISSN 1044-5498. doi:10.1089/dna.2020.5351.(англ.)
- Farajollahi, Sanaz and Maas, Stefan (2010). Molecular diversity through RNA editing: a balancing act. Trends in Genetics (Elsevier) 26 (5): 221–230.
- Costa Cruz, Pedro Henrique; Kato, Yuki; Nakahama, Taisuke; Shibuya, Toshiharu; Kawahara, Yukio (2020). A comparative analysis of ADAR mutant mice reveals site-specific regulation of RNA editing. RNA 26 (4): 454–469. ISSN 1355-8382. doi:10.1261/rna.072728.119.
- Fischer, Sylvia E. J.; Ruvkun, Gary (2020). Caenorhabditis elegans ADAR editing and the ERI-6/7/MOV10 RNAi pathway silence endogenous viral elements and LTR retrotransposons. Proceedings of the National Academy of Sciences 117 (11): 5987–5996. ISSN 0027-8424. doi:10.1073/pnas.1919028117.(англ.)
- Zhang, Jianzhi; Duan, Yuange; Dou, Shengqian; Luo, Shiqi; Zhang, Hong; Lu, Jian (2017). Adaptation of A-to-I RNA editing in Drosophila. PLOS Genetics 13 (3): e1006648. ISSN 1553-7404. doi:10.1371/journal.pgen.1006648.
- Zhou, Shili; Yang, Chao; Zhao, Fanfan; Huang, Yanxia; Lin, Yuxia; Huang, Changbai; Ma, Xiaocao; Du, Jingjie; Wang, Yi; Long, Gang; He, Junfang; Liu, Chao; Zhang, Ping (2019). Double-stranded RNA deaminase ADAR1 promotes the Zika virus replication by inhibiting the activation of protein kinase PKR. Journal of Biological Chemistry 294 (48): 18168–18180. ISSN 00219258. doi:10.1074/jbc.RA119.009113.
- Moroz, Leonid L.; Kocot, Kevin M.; Citarella, Mathew R.; Dosung, Sohn; Norekian, Tigran P.; Povolotskaya, Inna S.; Grigorenko, Anastasia P.; Dailey, Christopher; Berezikov, Eugene; Buckley, Katherine M.; Ptitsyn, Andrey; Reshetov, Denis; Mukherjee, Krishanu; Moroz, Tatiana P.; Bobkova, Yelena; Yu, Fahong; Kapitonov, Vladimir V.; Jurka, Jerzy; Bobkov, Yuri V.; Swore, Joshua J.; Girardo, David O.; Fodor, Alexander; Gusev, Fedor; Sanford, Rachel; Bruders, Rebecca; Kittler, Ellen; Mills, Claudia E.; Rast, Jonathan P.; Derelle, Romain; Solovyev, Victor V.; Kondrashov, Fyodor A.; Swalla, Billie J.; Sweedler, Jonathan V.; Rogaev, Evgeny I.; Halanych, Kenneth M.; Kohn, Andrea B. (2014). The ctenophore genome and the evolutionary origins of neural systems. Nature 510 (7503): 109–114. ISSN 0028-0836. doi:10.1038/nature13400.
- Barak, Michal; Porath, Hagit T.; Finkelstein, Gilad; Knisbacher, Binyamin A.; Buchumenski, Ilana; Roth, Shalom Hillel; Levanon, Erez Y.; Eisenberg, Eli (2020). Purifying selection of long dsRNA is the first line of defense against false activation of innate immunity. Genome Biology 21 (1). ISSN 1474-760X. doi:10.1186/s13059-020-1937-3.(англ.)
- Nakahama, Taisuke; Kawahara, Yukio (2020). Adenosine-to-inosine RNA editing in the immune system: friend or foe?. Cellular and Molecular Life Sciences. ISSN 1420-682X. doi:10.1007/s00018-020-03466-2.(англ.)
- Khermesh, Khen; D'Erchia, Anna Maria; Barak, Michal; Annese, Anita; Wachtel, Chaim; Levanon, Erez Y.; Picardi, Ernesto; Eisenberg, Eli (2016). Reduced levels of protein recoding by A-to-I RNA editing in Alzheimer's disease. RNA 22 (2): 290–302. ISSN 1355-8382. doi:10.1261/rna.054627.115.
- Gaisler-Salomon, Inna; Kravitz, Efrat; Feiler, Yulia; Safran, Michal; Biegon, Anat; Amariglio, Ninette; Rechavi, Gideon (2014). Hippocampus-specific deficiency in RNA editing of GluA2 in Alzheimer's disease. Neurobiology of Aging 35 (8): 1785–1791. ISSN 01974580. doi:10.1016/j.neurobiolaging.2014.02.018.
- Yang, Weidong; Wang, Qingde; Kanes, Stephen J; Murray, John M; Nishikura, Kazuko (2004). Altered RNA editing of serotonin 5-HT2C receptor induced by interferon: implications for depression associated with cytokine therapy. Molecular Brain Research 124 (1): 70–78. ISSN 0169328X. doi:10.1016/j.molbrainres.2004.02.010.
- Samuel, Charles E. (2011). Adenosine deaminases acting on RNA (ADARs) are both antiviral and proviral. Virology 411 (2): 180–193. ISSN 00426822. doi:10.1016/j.virol.2010.12.004.
- Priyadharsini, Jayaseelan Vijayashree; Paramasivam, Arumugam (2021). RNA editors: key regulators of viral response in cancer patients. Epigenomics. ISSN 1750-1911. doi:10.2217/epi-2021-0001.
- Piontkivska, Helen; Matos, Luis F.; Paul, Sinu; Scharfenberg, Brian; Farmerie, William G.; Miyamoto, Michael M.; Wayne, Marta L. (2016). Role of Host-Driven Mutagenesis in Determining Genome Evolution of Sigma Virus (DMelSV; Rhabdoviridae) inDrosophila melanogaster. Genome Biology and Evolution 8 (9): 2952–2963. ISSN 1759-6653. doi:10.1093/gbe/evw212.
- Doria, Margherita; Neri, Francesca; Gallo, Angela; Farace, Maria Giulia; Michienzi, Alessandro (2009). Editing of HIV-1 RNA by the double-stranded RNA deaminase ADAR1 stimulates viral infection. Nucleic Acids Research 37 (17): 5848–5858. ISSN 1362-4962. doi:10.1093/nar/gkp604.
- Montiel-Gonzalez, Maria Fernanda; Diaz Quiroz, Juan Felipe; Rosenthal, Joshua J.C. (2019). Current strategies for Site-Directed RNA Editing using ADARs. Methods 156: 16–24. ISSN 10462023. doi:10.1016/j.ymeth.2018.11.016.
- Qu, Liang; Yi, Zongyi; Zhu, Shiyou; Wang, Chunhui; Cao, Zhongzheng; Zhou, Zhuo; Yuan, Pengfei; Yu, Ying; Tian, Feng; Liu, Zhiheng; Bao, Ying; Zhao, Yanxia; Wei, Wensheng (2019). Programmable RNA editing by recruiting endogenous ADAR using engineered RNAs. Nature Biotechnology 37 (9): 1059–1069. ISSN 1087-0156. doi:10.1038/s41587-019-0178-z.
Джерела
- Farajollahi, Sanaz and Maas, Stefan (2010). Molecular diversity through RNA editing: a balancing act. Trends in Genetics (Elsevier) 26 (5): 221–230.