Кровотворення
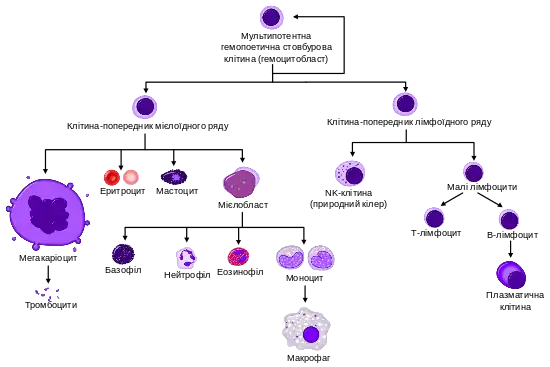
Кровотво́рення (лат. haemopoesis), гематопое́з або гемопое́з – процес утворення клітин крові із гемопоетичних стовбурових клітин (інша назва – гемоцитобласти). Гемоцитобласти дають початок клітинам-попередникам, які інтенсивно діляться і поступово диференціюються у зрілі формені елементи крові. На перших етапах кровотворення розгалужується на дві лінії: мієлоїдну (дає початок еритроцитам, мегакаріоцитам, гранулоцитам та моноцитам) та лімфоїдну (дає початок В-лімфоцитам, Т-лімфоцитам та природним кілерам (NK-клітинам). В нормі в організмі людини утворюється приблизно 1011-1012 нових формених елементів на добу, для підтримання їх постійного рівня.
Місця кровотворення
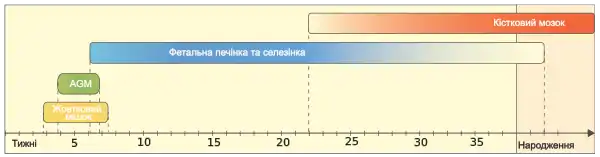
В організмі дорослої людини єдиним органом кровотворення є кістковий мозок. Проте, під час ембріонального розвитку гемопоез починається задовго до появи в зародка кісток. Перші етапи кровотворення відбуваються у жовтковому мішку, такий гемопоез називають «примітивним», його основне завдання – утворення еритроцитів, що сприятимуть кращому газообміну у тканинах зародка, який швидко росте. Інші клітини крові ще не утворюються. Еритроцити, на цій стадії містять ембріональний гемоглобін (HbE). У жовтковому мішку кровотворення тісно пов'язане із утворенням клітин ендотелію судин, вважається, що червоні кров’яні тільця та ендотеліоцити походять із спільного попередника — стовбурових клітин гемангіобластів. [1]. Примітивний гемопоез у людини триває до кінця другого місяця ембріонального розвитку [2].
У ссавців наступним місцем кровотворення є ділянка, що оточує дорзальну аорту, її називають AGM (від англ. aorta-gonad-mesonephros, аорта-гонади-мезонефрос), тут починається дефінітивний (остаточний) гемопоез. У людини гемопоез в ділянці дорзальної аорти триває від 27 до 40 дня ембіріогенезу [3]. Незабаром гемопоетичні стовбурові клітини мігрують у печінку та селезінку зародка, які є основними місцями кровотворення від 6 тижня до 6-7 місяця ембіріонального розвитку, продукування клітин крові на низькому рівні тут продовжується до 2-ох тижнів після народження [2]. Кістковий мозок стає основним місцем геомопоезу на 6-7 місяць розвитку зародка, а до кінця першого місяця постнатального періоду залишається єдиним місцем, в якому утворюються нові формені елементи крові.
У ранньому дитинстві весь кістковий мозок залучений у гемопоезі, проте згодом у довгих кістках гемопоетичні клітини починаються замінюватись жировою тканиною. У дорослих людей червоний кістковий мозок, що забезпечує кровотворення зберігається в епіфізах довгих кісток та у плоских кістках, навіть тут він на 50% складається із жиру. Інколи у дорослих людей кровотворення може відновлюватись у печінці та селезінці, такий стан називається екстрамедулярним гемопоезом, і є патологічним [2].
Гемопоетичні стовбурові клітини
Кровотворення розпочинається із гемопоетичних стовбурових клітин (ГСК) або гемоцитобластів. В організмі людини таких клітин дуже мало, зазвичай менше ніж одна ГСК на 5×104 клітин кісткового мозку [4] (за іншими джерелами – 1 ГСК на 2×107[2]). Стовбурові клітини мають дві основні властивості: здатність диференціюватись у інші типи клітин та спроможність підтримувати власну популяцію на сталому рівні.
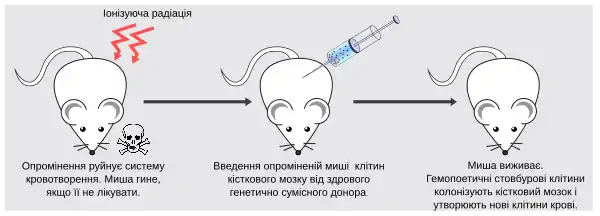
Завдяки самовідновленню кількість гемопоетичних стовбурових клітин залишається приблизно сталою впродовж всього життя особини. Проте, коли організм потребує швидких темпів кровотворення, ГСК можуть демонструвати величезну проліферативну активність. Ця їхня властивість може бути продемонстрована у досліді із мишами, яким повністю руйнують систему кровотворення, шляхом опромінення летальною дозою радіації (950 рад). Опромінені миші помирають за 10 днів, якщо їм не пересадити нормальні клітини кісткового мозку від генетично сумісного донора. Хоча нормальна миша має 3×108 клітин кісткового мозку, введення тільки 104-105 донорських клітин (тобто 0,01-0,1% від нормальної кількості) є достатнім для повного відновлення системи кровотворення, що доводить величезну здатність стовбурових клітин до проліферації та диференціації [4].
У схожому досліді була доведена мультипотентність ГСК: клітини донора перед введенням в організм опроміненої миші мітились спеціальними генетичними маркерами. Такі маркери можуть вводитись у клітини за допомогою ретровірусних векторів (як і всі ретровіруси вони можуть вбудовувати свій геном у геном клітини, але не здатні до утворення вірусного потомства). Через деякий час після введення мічених клітин миші, її кров'яні тільця перевіряються на наявність відповідної генетичної мітки. Оскільки всі формені елементи крові у миші-реципієнта були міченими, це свідчить про, те що вони є нащадками мультипотентних стовбурових клітин, введених від донора [5].
Маркери диференціації гемопоетичних стовбурових клітин
Маркери (або кластери) диференціації – це молекули, які наявні тільки на поверхні одного або кількох видів клітин. Використання флюоресцентно мічених антитіл до маркерів диференціації дозволяє отримувати майже чисті препарати одного типу клітин за допомогою методу протокова цитометрії, а саме флуоресцентно-активованого сортування клітин (англ. fluorescence-activated cell sorting (FACS)).
Перші спроби виділити популяцію гемопоетичних стовбурових клітин у 1988 році здійснив Вайсман (англ. I.L.Weissman) та колеги. Їхній підхід базувався на негативній селекції: оскільки маркери диференціації для ГСК були невідомі, але відомі для інших типів клітин, то із препаратів кісткового мозку за допомогою флуоресцентно-активованого сортування видалялись всі зрілі клітини крові. Після цього клітини, що залишились тестувались на здатність відновлювати кровотворення в опромінених мишей. Використання негативної селекції дозволило досягнути такого рівня концентрації ГСК, за якого всього тридцяти клітин із очищеного препарату було достатньо для відновлення гемопоезу [6]. Також було з’ясовано, що на поверхні гемоцитобластів мишей наявний поверхневий антиген Sca-1 (stem-cell antigen 1).
Пізніше був відкритий маркер диференціації CD34, наявний на поверхні невеликої популяції (близько 1 %) людських клітин залучених до кровотворення, яка охоплює і стовбурові клітини. Відкриття цього маркера дозволило зробити процедуру виділення ГСК досить простою і досягати рівнів очищення, за яких навіть одна клітина із певною вірогідністю може відновити всю систему кровотворення [4].
Відомі кластери диференціації для гемопоетичних стовбурових клітин мишей та людей наведені у таблиці.
| Таблиця маркерів диференціації геомопоетичних ствобурових клітин [7] | |
| Миша | Людина |
| CD34low/- | CD34+ |
| Sca1+ | CD59+ 1 |
| Thy1+/low | Thy1+ |
| CD38+ | CD38low/- |
| C-kit+ | C-kit-/low |
| lin- 2 | lin- 2 |
| 1 Була оцінена тільки одна родина CD59 маркерів
2 На lin- клітинах відсутні від 13 до 14 макреів диференціювання зрілих клітин крові | |
Моделі для вивчення гемопоетичних стовбурових клітин людини
Основною моделлю для виявлення та дослідження властивостей людських ГСК є миші із важким комбінованим імунодефіцитом (англ. severe combined immunodeficiency, SCID). В них не працює специфічна імунна система (відсутні Т і В лімфоцити), що бере участь у відторгненні чужорідних клітин, тканин і органів. Через це людські тканини кісткового мозку, що містять ГСК, а також тканини тимуса добре приживаються у мишей із SCID. Після введення дослідним тваринам різних субпопуляцій людських CD34+ клітин із кісткового мозку, досліджується розвиток клітин крові людини. За відсутності людських факторів росту розвивається тільки невелика кількість клітин-попередників гранулоцитів та макрофагів. Проте, якщо в організм миші ввести разом із популяцією CD34+ еритропоетин та інші цитокіни, спостерігається утворення клітин-попередників та зрілих клітин мієлоїдного, лімфоїдного та еритроїдного рядів[4].
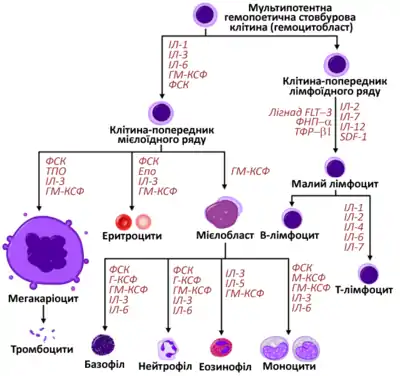
Комітовані клітини-попередники
_diagram_uk.png.webp)
На перших етапах кровотворення, мультипотентні стовбурові клітини можуть диференціюватись по одному із двох шляхів, даючи початок або клітині-попереднику лімфоїдного ряду, або клітині-попереднику мієлоїдного ряду. На цей процес впливає мікрооточення, зокрема наявність та концентрація певних факторів росту. Клітини-попередники лімфоїдного чи мієлоїдного ряду відрізняються від ГСК своєю нездатністю до самовідновлення, а також тим, що вони комітовані до певного клітинного ряду (тобто їхня доля вже визначена):
- Комітована клітина-попередник лімфоїдного ряду дає початок Т і В лімфоцитам, а також NK-клітинам (англ. natural killer cell) та деяким дендритним клітинам;
- Комітована клітина попередник мієлоїдного ряду дає початок іншим клітинам крові – еритроцитам (червоним кров'яним тільцям), більшості лейкоцитів (нейтрофілам, еозинофілам, базофілам, моноцитам, мастоцитам та дендритним клітинам) та мегакаріоцитам (від яких відшнуровуються тромбоцити)[4].
Комітування є поступовим процесом. До того, як клітина-попередник перестане проліферувати і стане зрілою, вона встигає багато разів поділитись і дати початок великій кількості відповідних спеціалізованих клітин. Через це, внаслідок всього лише одного поділу ГСК, може утворитись кілька тисяч зрілих клітин різних типів. Саме тому ГСК становлять тільки дуже невеличку частину від загальної популяції клітин кісткового мозку. Із цієї ж причини можливо підтримувати високі темпи кровотворення навіть за умов дуже повільного поділу стовбурових клітин. А це важливо для того, щоби зменшити ризик виникнення мутацій у стовбурових клітинах, яке могло би мати наслідком постійне утворення колоній мутантних клітин в організмі. Повільний поділ ГСК також потрібний, щоб уникнути реплікативного старіння. Гемоцитобласти, модифіковані таким чином, що вони діляться швидко (наприклад шляхом нокауту гену Gfi 1, який обмежує швидкість проліферації) не можуть забезпечувати кровотворення впродовж всієї тривалості життя організму[5].
Роль клітин строми у кровотворенні
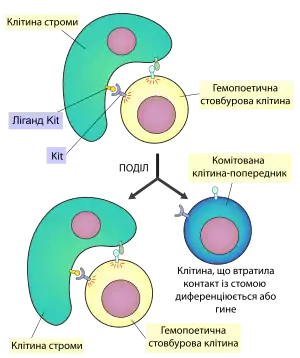
Крім кровотвірних клітин до кісткового мозку входять також і клітини строми (остеобласти, жирові клітини, ендотеліальні клітини, фібробласти та макрофаги), що утворюють гемопоез-індукуюче мікрооточення, потрібне для росту та диференціації ГСК і клітин-попередників. Гемопоез-індукуюче оточення складається із власне клітинного матриксу та гемопоетичних ростових факторів. Більшість із цих факторів є розчинними агентами, що досягають своєї мішені шляхом дифузії, інші ж можуть бути мембранозв'язаними і для реалізації свого ефекту потребувати безпосереднього клітинного контакту [4].
Клітини строми, зокрема остеобласти, також потрібні для підтримання гемоцитобластами статусу недиференційованих стовбурових клітин. Гемопоетичні стовбурові клітини можуть існувати у кістковому мозку тільки в окремих нішах, що створюються стромою. Основна властивість цієї ніші полягає у тому, що мікрооточення стимулює в ГСК сигнальний шлях Wnt (шлях, необхідний для підтримання стовбуровості). Також були виявлені інші сигнальні шляхи, задіяні в цьому процесі. Зокрема у мишей із специфічним синдромом, що включав анемію внаслідок нестачі еритроцитів, стерильність через відсутність гамет та нестачу пігментних клітин, спостерігались мутації у одному із двох генів: гені рецепторної тирозинкінази Kit або її ліганду. Було встановлено, що ліганд Kit експресується на поверхні стромальних клітин, і повинен бути мембранозв'язаним для того, щоб виконувати свою роль у підтриманні популяції стовбурових клітин. Інша назва цього білка - фактор стовбурових клітин (англ. Stem cell factor, SCF), він також може існувати і в розчинній формі, діючи не тільки не гемоцитобласти, а й на більш диференційовані клітини [8]. Рецепторна тирозин-кіназа Kit присутня на поверхні стовбурових клітин (зокрема ГСК)[5].
Роль цитокінів у кровотворенні
Всі етапи кровотворення (підтримання популяції стовбурових клітин, проліферація, диференціація, апоптоз клітин різних ліній) залежать від наявності у середовищі розчинних або мембранозв'язаних ростових факторів. Різні кровотвірні клітини чутливі до різних цитокінів, власне, процес комітування великою мірою, і полягає у набутті чи втраті клітиною рецептора до певного цитокіну. Дія різних цитокінів часто перекривається, вони можуть бути синергістами або проявляти кумулятивний ефект на одну лінію кровотворення. З іншого боку, один і той же цитокін може впливати на кілька гілок гемопоезу. Кровотвірні фактори росту синтезуються як клітинами кісткового мозку, так і іншими тканинами організму (зрілими клітинами імунної системи, печінки і нирок). Вивчення впливу окремих ростових факторів на кровотворення можна проводити за допомогою культивування клітин кісткового мозку in vitro.
Вивчення значення цитокінів у кровотворенні за допомогою культур клітин кісткового мозку
Кровотвірні клітини кісткового мозку можливо вирощувати у культурі. Для цього стромальні клітини культивуються у моношарі в чашках Петрі, пізніше на цей моношар висівають свіжовиділені гемопоетичні клітини. В таких умовах вони можуть рости, ділитись і утворювати великі колонії. Якщо здійснювати культивування у напіврідкому агарі, то колонії будуть іммобілізовані, що дає змогу визначати типи клітин.
Для того щоб з'ясувати роль певного цитокіну у процесі кровотворення, його додають до культур клітин кісткового мозку і аналізують, які типи клітин утворюватимуться. До цитокінів, виявлених таким методом, належить сім'я кислих глікопротеїнів – колонієстимулюючих факторів (КСФ), названих так за їхню здатність індукувати формування окремих ліній кровотворних клітин[4].
Цитокіни, що впливають на еритропоез
Одним із перших виявлених гемопеотичних цитокінів був глікопротеїн еритропоетин, що виробляється нирками у відповідь на нестачу кисню або малу кількість еритроцитів в організмі. Оскільки рівень червоних кров'яних тілець зростає вже на 1-2 день після підвищення вмісту еритропоетину в крові, цей цитокін повинен діяти на безпосередні попередники еритроцитів. Клітини чутливі до еритропоетину можна виявити, додавши його до культури кісткового мозку у напіврідкому агарі. Через кілька днів з'являються невеликі колонії (приблизно по 60 еритроцитів), кожна із яких утворена однією комітованою клітиною-попередником еритроїдного ряду. Для поділу і виживання цих клітин необхідний еритропоетин. На більш ранні попередники цього ряду еритропоетин не діє, зате вони чутливі до іншого цитокіну - інтерлейкіну 3 (ІЛ-3). При додаванні цього фактора до культури кровотвірних клітин за 7-10 днів розвиваються колонії, що містять значно більшу кількість еритроцитів (приблизно 5000 у кожній колонії)[5].
Роль цитокінів у розвитку макрофагів та нейтрофілів
Два типи клітин, що здійснюють фагоцитоз — нейтрофіли та макрофаги — походять від спільного попередника (ГМ клітина-попередник). Було виявлено як мінімум сім різних колонієстимулюючих факторів, що викликають формування фагоцитів. Вважається, що діючи у різних комбінаціях, вони селективно регулюють продукування цих клітин in vivo. Ці КСФ утворюються різними типами клітин, зокрема ендотеліальними, фібробластами, макрофагами та лімфоцитами, їхні концентрації у крові здебільшого сильно збільшуються у відповідь на бактерійну інфекцію, внаслідок чого зростає і кількість фагоцитуючих клітин, що вивільняються у кровотік.
Інтерлейкін-3 є одним із найменш специфічних факторів, що впливає як на ГМ клітини-попередники, так і на гемоцитобласти і більшість інших класів комітованих клітин. Багато інших факторів проявляють більш селективну дію на лінію фагоцитуючих клітин. Всі ці КСФ, як і еритропоетин, є глікопротеїнами. Вони ефективні у низьких концентраціях (приблизно 10-12М) і діють через приєднання до специфічних клітинних рецепторів. Серед цих рецепторів є трансмембранні тирозинкінази, але більшість із них належать до великої родини цитокінових рецепторів. Члени цієї родини складаються із двох або більшої кількості субодиниць, одна із яких, зазвичай, є спільною для кількох типів рецепторів (наприклад для рецептора до ІЛ-3 та ГМ КСФ). КСФ, здебільшого, впливають не тільки на клітини-попередники, змушуючи їх продукувати більшу кількість зрілих клітин, а й на самі диференційовані клітини, активуючи їхні функції (наприклад фагоцитоз та знищення клітини-мішені). Клонування генів цих цитокінів дозволяє отримувати велику кількість білків, що стимулюють гемопоез в експериментальних тварин. Зараз такі білки широко використовуються у медицині для активізації розвитку кровотвірних тканин та підсилення стійкості до інфекцій.
Характеристика основних цитокінів залучених у кровотворення
| Основні цитокіни задіяні у кровотворенні [4][5] | |||
| Фактор | Основні функції | Клітини, які продукують | Родина рецепторів |
| Ліганд Kit (Фактор стовбурових клітин, англ. Stem cell factor) | Підтримання популяції стовбурових клітин, виживання і проліферація клітин-попередників, диференціація мастоцитів | Конститутивно експресується клітинами строми кісткового мозку | Рецепторні тирозинкінази |
| Еритропоетин | Утворення еритроцитів | Печінка, нирки | Цитокінова родина |
| Тромбопоетин | Утворення тромбоцитів | Печінка, нирки | Цитокінова родина [9] |
| КСФ гранулоцитів/макрофагів (ГМ КСФ) | Утворення гранулоцитів та макрофагів, активація і дозрівання дендритних клітин | Макрофаги, мастоцити, Т лімфоцити | Цитокінова родина |
| КСФ гранулоцитів | Утворення нейтрофілів та стимулювання їхньої дії | Фібробласти, макрофаги | Цитокінова родина |
| КСФ макрофагів | Утворення макрофагів та остеокластів, стимулювання їхньої дії | Фібробласти, макрофаги, ендотеліальні клітини | Рецепторні тирозин кінази |
| Ліганд Flt-3 | Розмноження ранніх клітин-попередників, утворення пре-В лімфоцитів | Фібробласти, ендотеліальні клітини | Рецепторні тирозинкінази[10] |
| Інтерлейкін 3 (ІЛ-3) | Проліферація ГСК та більшості клітин-попередників мієлоїдного ряду | Т лімфоцити, ендотеліальні клітини, макрофаги | Цитокінова родина |
| Інтерлейкін 5 (ІЛ-5) | Утворення еозинофілів | Активовані Т-гелпери | Цитокінова родина |
| Інтерлейкін 6 (ІЛ-6) | Стимулювання клітин-попередників, утворення тромбоцитів, продукція антитіл В-лімфоцитами | Активовані Т-лімфоцити, моноцити, фібробласти, ендотеліальні клітини | Цитокінова родина |
| Інтерлейкін 7 (ІЛ-7) | Продукування та виживання Т-лімфоцитів | Клітини строми кісткового мозку та тимуса | Цитокінова родина |
Генетичний контроль кровотворення
Розвиток мультипотентних гемопоетичних стовбурових клітин по одному із шляхів диференціації потребує експресії різних наборів генів у відповідний час і у правильному порядку. Регуляція експресії цих генів відбувається за участі транскрипційних факторів, які беруть безпосередню роль у програмуванні ліній гемопоезу. Частина із факторів транскрипції потрібна для процесу комітування клітин, інша - для синтезу білків, специфічних для даної лінії. Більшість знань про роль окремих генів у кровотворенні отримані шляхом їх нокауту (виключення).
Деякі із транскрипційних факторів активні у клітинах кількох ліній кровотворення, тоді експресія інших обмежена тільки однією. До поліфункціональних факторів належить GATA-2 — член родини транскрипційних факторів, що розпізнають тетрануклеотидну послідовність GATA у регуляторних ділянках генів-мішеней. Функціонування гену GATA-2 є необхідним для розвитку лімфоїдної, еритроїдної та мієлоїдної ліній. Як і слід очікувати, тварини із нокаутом цього гену гинуть під час ембріогенезу. На відміну від GATA-2, транскрипційний фактор Ikaros потрібний тільки для розвитку клітин лімфоїдного ряду. Хоча нокаутні по гену Ikaros миші не можуть утворювати достатньої кількості В-, Т- та NK-лімфоцитів, продукування еритроцитів, гранулоцитів та інших клітин мієлоїдного ряду у них не порушене. Такі тварини переживають ембріональний розвиток, але гинуть у перші дні після народження через важкий імунодефіцит[4].
Багато із транскрипційних факторів, які беруть участь у визначенні подальшої долі клітини, безпосередньо взаємодіють між собою. При чому ключові фактори певної лінії одночасно активують гени, потрібні для розвитку клітини по цій лінії, і пригнічують фактори, що сприяють іншому вибору. Прикладом антагоністичної дії транскрипційних факторів може бути взаємодія GATA-1 і PU.1 у клітині-попереднику мієлоїдного ряду. Ці білки фізично взаємодіють між собою, інгібуючи один одного, якщо переважає кількість білка GATA-1, то клітина буде розвиватись в еритроцит або тромбоцит, а якщо переважить PU.1 із неї в подальшому утвориться гранулоцит або моноцит. Схожим чином відбуваються і наступні кроки диференціації[1].
Спільною ознакою багатьох транскрипційних факторів, задіяних у кровотворенні людини, є те, що соматичні мутації або хромосомні транслокації, які зачіпають їхні гени, здебільшого призводять до злоякісного переродження клітин і розвитку різних форм лейкемії[1].
Апоптоз і кровотворення
Апоптоз – це запрограмована клітинна смерть, яка відбувається таким чином, щоб не завдати шкоди навколишнім тканинам, залишки клітини швидко поглинаються макрофагами. Апоптоз відіграє важливу роль у підтриманні постійної кількості кров'яних тілець, зокрема лейкоцитів. Кожен лейкоцит має визначений термін життя, після закінчення якого в ньому вмикаються механізми запрограмованої смерті. У дорослої людини в крові циркулює близько 5×1010 нейтрофілів, вони живуть всього кілька днів до того, як у них ініціюється апоптоз. Зрівноважені процеси загибелі і постійного утворення нових нейтрофілів підтримують їх постійний рівень у крові. Якщо апоптоз із якихось причин не може відбутись, це може призвести до виникнення лейкемії. Запрограмована клітинна смерть також допомагає підтримувати потрібну кількість комітованих клітин-попередників. Якщо на них не діють ростові фактори, клітини гинуть шляхом апоптозу.
У здійсненні апоптозу залучено велику кількість білків, частина яких відповідає за його стимуляцію, а інша частина — за пригнічення. До супресорів апоптозу, належать білки родини Bcl-2 (bcl-2 і bcl-XL). Bcl-2 відіграє важливу роль у регулюванні тривалості життя клітин різних ліній кровотворення, включно із лімфоцитами. Доросла людина в середньому має 5л крові із приблизно 2000 лімфоцитами на 1мм3 (загалом близько 1010 лімфоцитів). Під час гострої фази інфекції кількість лімфоцитів зростає від 4-ох до 15-ти разів. Оскільки імунна система не може підтримувати таку величезну кількість клітин впродовж тривалого часу, вона повинна позбутись їх частини після того як антигенна небезпека мине. Тому активовані лімфоцити мають нижчий рівень експресії Bcl-2, і, таким чином, є чутливішими до індукції апоптозу, ніж неактивовані лімфоцити і клітини імунної пам'яті. Проте, якщо лімфоцит продовжує стимулюватись антигеном, цей сигнал блокуватиме апоптоз. Після зниження рівня антигенів, знижується і рівень блокування апоптозу, і лімфоцити гинуть.
Стохастичні процеси під час кровотворення
Дослідження in vitro показують, що у поведінці гемопоетичних стовбурових клітин велику роль відіграє випадок — відображення «шумів» у системі контролю експресії генів. Якщо дві сестринські клітини-попередники розділити одразу ж після мітозу і культивувати у максимально ідентичних умовах (в тому числі й у присутності однакових концентрацій колонієзаохочувальних чинників), вони дають початок колоніям, що відрізняються за типами і розміром клітин та їх співвідношеннями. Схожі результати спостерігаються і при культивуванні клітин, що були спеціально селекціоновані на максимальну схожість між собою.
Таким чином, як програмування клітинних поділів, так і процес комітування до відповідної лінії диференціації певною мірою включає випадкові події на рівні окремих клітин, попри те, що регуляція функцій цілого організму контролюється більш точними методами. Колонієстимулюючі фактори діють на кровотвірні клітини не прямо «диктуючи» клітині, яким шляхом вона повинна розвиватись, а тільки змінюючи імовірність тої чи іншої поведінки[5].
Гомеостаз кровотворення
Гомеостаз кровотворення – це процес підтримання сталого рівня формених елементів крові, під час якого кількість новоутворених клітин крові рівна кількості втрачених. Середня тривалість життя еритроцита становить 120 днів, перед тим як він буде фагоцитований і перетравлений макрофагами у селезінці. Тривалість життя різних лейкоцитів коливається від кількох днів для нейтрофілів до 20-30 років для деяких Т-лімфоцитів.
Кровотворення контролюється складними механізмами, що забезпечують постійну кількість кожного типу клітин крові. Водночас ці механізми достатньо гнучкі, що дозволяє збільшувати темпи гемопоезу у 10-20 разів у випадку крововтрати або інфекції. До того-ж кожен тип інфекції по-різному впливає на кровотворення, наприклад деякі бактерійні інфекції викликають селективне збільшення продукування нейтрофілів, тоді як у відповідь на протозойне зараження зростає рівень еозинофілів. Через це лейкоцитарну формулу крові можна використовувати для діагностики інфекційних та інших запальних захворювань. За певних умов може відбуватись також і селективне збільшення кількості еритроцитів, наприклад при акліматизації до життя на великих висотах.
До механізмів регуляції гомеостазу кровотворення належать:
- Контроль рівню продукції цитокінів клітинами строми кісткового мозку;
- Утворення цитокінів іншими типами клітин, наприклад активованими Т-лімфоцитами та макрофагами;
- Регуляція експресії рецепторів до гемопоетичних цитокінів у ГСК та комітованих клітинах-попередниках;
- Видалення деяких клітин за допомогою індукції апоптозу.
Порушення одного або декількох із цих регуляторних механізмів може призвести до серйозних і навіть летальних порушень функціонування організму. Наприклад, патологічні зміни в експресії певних гемопоетичних цитокінів та їхніх рецепторів має наслідком нерегульовану проліферацію клітин і виникнення деяких типів лейкімії.
Список використаної літератури
- Orkin SH, Zon LI (2008). Hematopoiesis: An Evolving Paradigm for Stem Cell Biology. Cell 132: 631–44. PMID 18295580. doi:10.1016/j.cell.2008.01.025.
- Hoffbrand V, Moss P, Pettit J (2006). Essential Haematology (вид. 5th). Wiley-Blackwell. ISBN 9781405136495. Архів оригіналу за 3 вересня 2011. Процитовано 10 червня 2011.
- Tavian M, Biasch K, Sinka L, Vallet J, Péault B (2010). Embryonic origin of human hematopoiesis. Int J Dev Biol 54: 1061–5. PMID 20711983. doi:10.1387/ijdb.103097mt.
- Kindt TJ, Osborne BA, Goldsby RA (2006). Kuby Immunology (вид. 6th). W. H. Freeman. ISBN 978-1429202114. Архів оригіналу за 8 березня 2012. Процитовано 10 червня 2011.
- Alberts B, Johnson A, Lewis J, Raff M, Roberts K, Walter P (2007). Molecular Biology of the Cell (вид. 5th). Garland Science. ISBN 978-0-8153-4105-5.
- Spangrude GJ, Heimfeld S, Weissman IL (1988). Purification and characterization of mouse hematopoietic stem cells. Science 241: 58–62. PMID 2898810. doi:10.1126/science.2898810.
- Stem cell information Архівовано 25 травня 2011 у Wayback Machine. від The National Institutes of Health resource for stem cell research
- McNiece IK, Briddell RA (1995). Stem cell factor. Journal of Leukocyte Biology 58: 14–22. PMID 7542304.
- Zhang Q, Pan RM, Ge YC, Xu P (2004). Expression of the soluble extracellular domain of human thrombopoietin receptor using a maltose-binding protein-affinity fusion system. Biol Pharm Bull. 27: 219–21. PMID 14758037.
- Gilliland DG, Griffin JD (2002). The roles of FLT3 in hematopoiesis and leukemia. Blood 100: 1532–42. PMID 12176867.
