Рибонуклеїнова кислота
Рибонуклеїнова кислота, РНК — клас нуклеїнових кислот, лінійних полімерів нуклеотидів, до складу яких входять залишок фосфорної кислоти, рибоза (на відміну від ДНК, що містить дезоксирибозу) і азотисті основи — аденін, цитозин, гуанін і урацил (на відміну від ДНК, що замість урацила містить тимін).
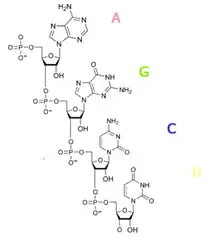
Рибонуклеїнова кислота — одна з трьох основних макромолекул (дві інші ДНК та білки), яка грає важливу роль в кодуванні, зчитуванні, регулюванні та вираженні генів. Так само як ДНК, РНК складається з ланцюжка нуклеотидів. Кожен нуклеотид складається з азотистої основи, цукру рибози та фосфатної групи. Послідовність нуклеотидів дозволяє РНК кодувати генетичну інформацію. Всі клітинні організми (еукаріоти) використовують РНК (мРНК) для програмування синтезу білків.
Клітинні РНК утворюються під час процесу, який називається транскрипцією, тобто синтезу РНК на матриці ДНК за допомогою спеціальних ферментів — РНК-полімерази. Після цього матричні РНК (мРНК) беруть участь в процесі, який називається трансляцією. Трансляція — це синтез білку на матриці мРНК за участі рибосом. Інші РНК після транскрипції піддаються хімічним модифікаціям, і, після утворення вторинних і третинних структур, виконують функції залежно від типу РНК.
Для одноланцюгових РНК характерні різноманітні просторові структури, в яких частина нуклеотидів одного і того ж ланцюга спарені між собою. Деякі високоструктуровані РНК беруть участь в синтезі білка клітини, наприклад транспортні РНК (тРНК) слугують для розпізнання кодонів і доставки відповідних амінокислот до місця синтезу білка, а рибосомні РНК слугують структурною і каталітичною основою рибосом.
Проте функції РНК в клітинах не обмежуються роллю в трансляції. Так, малі ядерні РНК беруть участь в сплайсингу еукаріотичних матричних РНК та інших процесах.
Окрім того, що молекули РНК входять до складу деяких ферментів (наприклад теломерази), в окремих РНК виявлена власна ферментативна активність: можливість вносити розриви в інші молекули РНК або, навпаки, «склеювати» два РНК фрагменти. Такі РНК називаються рибозимами.
РНК синтезуються в клітинах всіх клітинних живих організмів, а також містяться в віроїдах та деяких вірусах. Основні функції РНК в клітинних організмах залежать від типу РНК. Кодуючі РНК є матрицею для трансляції генетичної інформації в білки, некодуючі РНК виконують додаткові функції, такі як транспорт амінокислот до рибосом, регуляція експресії генів тощо. У вірусах РНК може бути носієм генетичної інформації, замість ДНК. Віроїди складаються з кільцевої молекули РНК та не містять в собі інших молекул.
Існує гіпотеза світу РНК, згідно з якою РНК виникли до білків й були першими формами життя.
Хімічна будова
Рибонуклеїнові кислоти як і інші нуклеїнові кислоти складаються з нуклеотидів — складних хімічних сполук, що утворені з моносахариду, азотистої основи пурину чи піримідину, залишків фосфорної кислоти. Моносахаридом в усіх РНК є рибоза — п'ятивуглецева молекула. До її першого (1') атому вуглецю за допомогою фосфодіефірного зв'язку приєднано азотисту основу, рибоза з такою основою утворює рибонуклеозид.
Основними нуклеозидами в природних біологічних полімерних РНК є аденозин, гуанозин, цитидин, а також уридин (на відміну від ДНК, де замість останнього присутній тимідин). Серед інших, мінорних РНК описано понад 100 інших нуклеозидів, серед яких найбільше поширені інозин, псевдоуридин, 5-метилцитидин тощо. Також трапляються РНК з модифікацією рибози, зокрема 2'-О-метилрибоза.
У клітинах поширені мономери рибонуклеїнової кислоти — рибонуклеотиди. Вони використовуються для синтезу полімерних РНК. Також аденінвмісні нуклеотиди АТФ і АДФ беруть участь у метаболізмі клітини, циклічний АМФ і гуанінвмісні ГТФ, ГДФ та цГМФ є сигнальними молекулами. Значно більше в клітинах полімерних РНК. Ці молекули утворюються шляхом з'єднання 5' і 3' OH-груп через молекулу фосфорної кислоти. Фосфатні групи при фізіологічному рН негативно заряджені, тому РНК — поліаніони, негативно заряджені полімери.
Полімерні молекули РНК переважно одноланцюгові, але в клітинах трапляються й дволанцюгові РНК: як вірусного походження, так і внутрішньоклітинного (мікроРНК, піРНК тощо).
Структура та топологія
Азотисті основи у складі РНК можуть утворювати водневі зв'язки між цитозином і гуаніном, аденіном і урацилом, а також між гуаніном і урацилом. Однак можливі й інші взаємодії, наприклад, кілька аденінів можуть утворювати петлю, або петля, що складається з чотирьох нуклеотидів, в якій є пара основ аденін — гуанін.
Важлива структурна особливість РНК, що відрізняє її від ДНК — наявність гідроксильної групи в 2' положенні рибози, яка дозволяє молекулі РНК існувати в А, а не В-конформації, що найчастіше спостерігається у ДНК. У А-форми глибока і вузька велика борозенка і неглибока і широка мала борозенка. Другий наслідок наявності 2' гідроксильної групи полягає в тому, що конформаційної пластичні, тобто не беруть участь в утворенні подвійної спіралі, ділянки молекули РНК можуть хімічно атакувати інші фосфатні зв'язки та їх розщеплювати.
«Робоча» форма одноланцюжкові молекули РНК, як і у білків, часто має третинну структурою. Третинна структура утворюється на основі елементів вторинної структури, що утворюється за допомогою водневих зв'язків усередині однієї молекули. Розрізняють декілька типів елементів вторинної структури — стебло-петлі, петлі і псевдовузли. У силу великої кількості можливих варіантів спарювання азотистих основ передбачення вторинної структури РНК — набагато складніше завдання, ніж передбачення вторинної структури білків, але в наш час[коли?] є ефективні програми, наприклад, mfold.
Прикладом залежності функцій молекул РНК від їх вторинної структури є ділянка внутрішньої посадки рибосоми (англ. IRES). IRES — структура на 5' кінці інформаційної РНК, яка забезпечує приєднання рибосоми в обхід звичайного механізму ініціації синтезу білка, що вимагає наявності особливого модифікованого нуклеотида (кепа) на 5' кінці молекули та додаткових білкових факторів ініціації. Спочатку IRES були виявлені у вірусних РНК, але зараз накопичується все більше даних про те, що клітинні мРНК також використовують IRES-залежний механізм ініціації в умовах стресу.[1]
Порівняння з ДНК
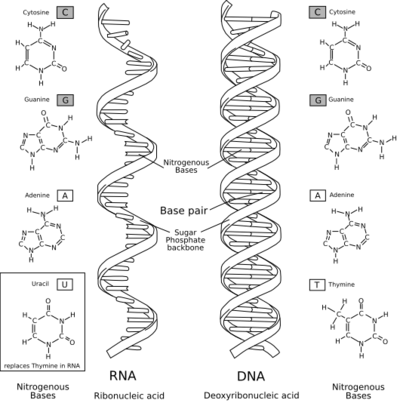
Між ДНК і РНК існують три основні відмінності: ДНК містить цукор дезоксирибозу, РНК — рибозу, у якої є додаткова, порівняно з дезоксирибозою, гідроксильна група. Ця група збільшує ймовірність гідролізу молекули, тобто зменшує стабільність молекули РНК.
Азотиста основа, компліментарна аденіну, у РНК не тимін, як у ДНК, а урацил — неметильована форма тиміну. ДНК існує у формі подвійної спіралі, що складається з двох окремих молекул. Молекули РНК, в середньому, набагато коротше і переважно одноланцюжкові.
Структурний аналіз біологічно активних молекул РНК, включаючи тРНК, рРНК, мяРНК та інші молекули, які не кодують білків, показав, що вони складаються не з однієї довгої спіралі, а з численних коротких спіралей, розташованих близько одна до одної і утворюють щось, схоже на третинну структуру білка. У результаті цього РНК може каталізувати хімічні реакції, наприклад, пептид-трансферазний центр рибосоми, що бере участь в утворенні пептидного зв'язку білків, повністю складається з РНК.
Взаємодія з білками
Більшість молекул РНК, наприклад, рРНК і мяРНК (малі ядерні РНК) в клітині функціонують у вигляді комплексів з білками.
Деякі РНК асоціюють з молекулами РНК після їх синтезу або (в еукаріотів) експорту з ядра в цитоплазму. Такі РНК-білкові комплекси називаються рибонуклеопротеїновими комплексами або рибонуклеопротеїдами. Серед найважливіших рибонуклеопротеїдів — рибосоми та сплайсосоми. Рибонуклеопротеїновий комлекс являє собою й теломераза тварин.
Чимало білків, які не пов'язані з РНК постійно, тим не менш мають РНК-зв'язувальні домени.
У функціонуванні РНК задіяні численні ферменти. Серед них РНК-полімерази, РНК-гелікази, нуклеази, лігази тощо. Окрема група ферментів редагування РНК модифікує окремі нуклеотиди, змінюючи генетичну інформацію. У тварин це аденозиндезамінази РНК, а в хребетних ще й цитидиндезамінази РНК. У рослин наявні спеціальні PPAR-білки.
Біологічні властивості
Синтез полімерної РНК здійснюється на матриці ДНК в ході транскрипції. Далі первинний транскрипт проходить модифікації: видаляються одні фрагменти (сплайсинг), додаються інші (наприклад, поліаденилювання), замінюються нуклеотиди (редагування РНК) тощо. Ці процеси називають процесингом РНК. Дозріла РНК далі транспортується до місця її функціонування. Особливо розвинений транспорт РНК у клітинах евкаріот. Існує як транспорт РНК з ядра до цитоплазми, так і транспорт по цитоплазмі великих клітин (наприклад, нейронів).
Усі молекули РНК мають певний час життя, а потім руйнуються механізмами деградації РНК, насамперед РНК-азами.
Процесинг
Чимало молекул РНК після синтезу на матриці ДНК підлягають змінам, які призводять до дозрівання новосинтезованої молекули до її функціонально активної форми. Ці процеси модифікації називають «процесингом», «котранскрипційною чи посттранскрипційною модифікацією».
Яскравим прикладом процесингу є дозрівання пре-мРНК до зрілої мРНК, з якої в цитоплазмі відбувається зчитування інформації про амінокислотну послідовність білків (трансляція). Інтрони вирізують з про-мРНК сплайсосомою, яка крім білків містить декілька малих ядерних РНК (мяРНК). Крім того, інтрони можуть каталізувати власне вирізання (аутосплайсинг). Утім, процесингу зазнають не лише мРНК, а й багато видів некодуючих РНК, транспортна РНК та рибосомна РНК.
У підручниках часто пишуть, що дозрівання пре-мРНК відбувається після її зчитування з ДНК матриці[2] . Таке явище відповідає лабораторним умовам (in vitro), коли стадії процесингу мРНК вивчають поступово, незалежно одна від одної. Але за умов in vivo, у живих клітинах, процесинг мРНК відбувається безпосередньо під час транскрипції в складі РНК-полімеразного комплексу[3]. Тому коректним терміном для мРНК дозрівання є котранскрипційна модифікація[2]. Проте не всі види РНК зазнають процесингу під час синтезу. Так, молекули тРНК дріжджів Saccharomyces cerevisiae дозрівають (зазнають сплайсингу) у цитоплазмі з подальшим експортом у ядро і поверненням до цитоплазми[4], тому в такому випадку коректним є термін «посттранскрипційна модифікація». Але у фаховій літературі частіше використовують термін «процесинг тРНК».
Біологічні функції та типи РНК
У сучасних клітинах РНК виконує функції, пов'язані з експресією генів та синтезом білків. Значна частина РНК задіяні в біосинтезі білків: транскрипції і трансляції генетичної інформації. Інші РНК регулюють ці процеси. Також молекули РНК беруть участь у процесах синтезу ДНК, зокрема при ініціації реплікації ДНК, подовженні теломерів, реакціях зворотної транскрипції.
Біосинтез білків
У біосинтезі білків задіяні матричні, рибосомні, транспортні РНК. Матричні РНК були дослідженні першими, тому їхня роль у кодуванні генетичної послідовності амінокислот у білку протиставлена всім іншим різноманітним РНК, які отримали назву «некодуючих РНК».
Матричні РНК
Матрична рибонуклеїнова кислота, або мРНК відповідає за перенесення інформації про первинну структуру білків від ДНК до місць синтезу білків. мРНК синтезується на основі ДНК в ході транскрипції, після чого, у свою чергу, використовується під час трансляції як матриця для синтезу білків. Тим самим мРНК грає важливу роль в «прояві» (експресії) генів.
Довжина типової зрілої мРНК становить від кількох сотень до кількох тисяч нуклеотидів. Найдовші мРНК відмічені у (+) ол-РНК-вмісних вірусів, наприклад пікорнавірусів, проте слід пам'ятати, що у цих вірусів мРНК утворює весь їхній геном.
ДНК нерідко порівнюють з кресленнями для виготовлення білків. Розвиваючи цю інженерно-виробничу аналогію, можна сказати, що, якщо ДНК — це повний набір креслень для виготовлення білків, що знаходиться на зберіганні в сейфі директора заводу, то мРНК — тимчасова робоча копія креслення, що видається в складальний цех.
Транспортні РНК
Транспортні РНК, або тРНК, є відносно малими молекулами, що складаються з приблизно 80 нуклеотидів. Вони мають характерну консервативну третинну структуру. Вони переносять специфічні амінокислоти до місця синтезу пептидного зв'язку в рибосомі. Кожна тРНК містить ділянку для приєднання амінокислоти і антикодон для пізнавання та приєднання до кодону мРНК. Антикодон утворює водневі зв'язки з кодоном, що поміщає тРНК в положення, що сприяє утворенню пептидного зв'язку між останньою амінокислотою утвореного пептиду і амінокислотою, приєднаною до тРНК.
тРНК приєднують певні амінокислоти в цитоплазмі і направляється до місця синтезу білка на мРНК де зв'язується з кодоном і віддає амінокислоту, яка використовується для синтезу білка.
Рибосомні РНК
Рибосомні РНК, або рРНК формують структуру рибосом. Еукаріотичні рибосоми містять чотири типи молекул рРНК: 18S, 5.8S, 28S і 5S. Три з чотирьох типів рРНК синтезуються в полісом. У цитоплазмі рибосомальні РНК з'єднуються з рибосомальними білками і формують нуклеопротеїн, званий рибосомою. Рибосома приєднується до мРНК і синтезує білок. рРНК становить до 80 % РНК, що виявляється в цитоплазмі еукаріотичної клітини.
Незвичайний тип РНК, який діє як тРНК і мРНК (тмРНК), виявлений у багатьох бактеріях і пластидах. При зупинці рибосоми на дефектних мРНК без стоп-кодонів тмРНК приєднує невеликий пептид, що направляє білок на деградацію.
Регуляція експресії генів
Низка малих дволанцюгових РНК виконують важливі функції з регуляції експресії генів. Зв'язуючись з мРНК або іншими РНК, вони призводять до їхньої деградації або принаймні зупинки трансляції. З'єднуючись з білками аргонавтами, ці молекули РНК утворюють комплекс RISC, який пригнічує трансляцію білка шляхом так званої РНК-інтерференції.
МікроРНК
МікроРНК — це малі РНК, довжиною 21-22 нуклеотиди наявні в еукаріот, які впливають через механізм РНК-інтерференції. При цьому комплекс мікроРНК і ферментів може призводити до метилювання нуклеотидів в ДНК промотора гена, що служить сигналом для зменшення активності гена. При використанні іншого типу регуляції мРНК, комплементарна мікроРНК, деградує. Однак є й мікроРНК, які збільшують, а не зменшують експресію генів.
Малі інтерферуючі РНК
Малі інтерферуючі РНК, або міРНК, подібний до мікроРНК клас молекул, довжиною 20—25 нуклеотидів. Часто утворюються в результаті розщеплення вірусних РНК, але існують і ендогенні клітинні міРНК. Малі інтерферуючі РНК також діють через РНК-інтерференцію за схожими з мікроРНК механізмами.
ПіРНК
ПіРНК — це малі інтерференційні РНК. Найбільша активність піРНК спостерігається в період ембріонального розвитку організмів, коли вони заглушують активність транспозонів.
мшРНК
Малі шпилькові РНК є штучними аналогами міРНК, розробленими з метою експериментального чи терапевтичного пригнічення експресії певних генів. Зазвичай вони є специфічними до певної послідовності та блокують трансляцію лише одного конкретного гену.
Еволюційна роль РНК
Молекули РНК присутні в більшості біологічних процесів. Це дало підстави думати про те, що на ранніх етапах біохімічної еволюції саме РНК була основним носієм генетичної інформації.
Оскільки мінорні азотисті основи РНК є не менш різноманітними ніж амінокислоти, низка дослідників, серед яких Крік і Спірін, висловили гіпотезу про існування на одному з етапів абіогенезу життя, побудованого на РНК. РНК могла виконувати роль як генетичної інформації (замість ДНК), як у сучасних РНК-вірусів, так і каталітичну (замість білків), що спостерігається в рибосомах та штучних рибозимах. Крім того, молекули РНК присутні в низці ключових процесів синтезу ДНК, що дозволяє вважати їх реліктовими механізмами з епохи, що передувала сучасному «світу ДНК».
РНК-віруси
У великої групи вірусів РНК відіграє роль носія генетичної інформації. При цьому в рослин існують інфекційні молекули РНК, віроїди, взагалі позбавлені білкової оболонки. У ретровірусів геном представлений РНК, але для експресії генів має утворитися компліментарна йому ДНК. У РНК-позитивних вірусів геном по суті є матричною РНК, тоді як у РНК-негативних потрібен синтез мРНК, компліментарної до РНК-геному. Також існують дволанцюгові РНК-віруси.
Рибозими
Рибозимами називають молекули РНК, здатні каталізувати певну реакцію. Це переважно штучно синтезовані невеликі молекули РНК, але по суті рибозимами є каталітичні центри рибосом.
«Реліктові» РНК
Реплікація ДНК у клітині неможлива без короткого олігонуклеотиду РНК - затравки, або праймеру. Його синтезує спеціальний фермент праймаза.
Інший олігонуклеотид РНК є кофактором ферменту теломерази, яка добудовує фрагменти ДНК на кінцях хромосом.
Використання людиною
РНК активно використовуються в наукових дослідженнях. Для аналізу РНК виділяють з тканин, окремих клітин, зовнішнього середовища. Існує низка фізико-хімічних методів, які дозволяють очистити РНК від інших речовин, зокрема ДНК.
Для прочитання послідовності РНК традиційно використовують метод зворотної транскрипції та подальшого аналізу кДНК. Натомість з 2010-х років активно розвиваються методи прямого прочитання - секвенування РНК. Після прочитання послідовностей РНК проводять їх біоінформатичний аналіз.
РНК вірусів досліджують задля виявлення їх активності, зокрема це стосується ВІЛ, коронавірусу SARS-Cov2 тощо. Деякі хвороби людини пов'язані зі зміною кількості певної РНК, що необхідно виміряти для діагностики.
У 2020 році у клінічному використанні з'явилися перші мРНК-вакцини, які містять генетичний код вірусного білку та виробляють його в організмі людини задля безпечного набуття імунітету без контакту з самими вірусом.
Модифіковані молекули РНК використовують для блокування надмірної експресії певного гену, як наприклад препарат патісіран.
Історія вивчення
Нуклеїнові кислоти відкрив в 1868 році швейцарський учений Іоганн Фрідріхм Мішер, який назвав ці речовини «нуклеїн», оскільки вони були виявлені в ядрі (лат. nucleus)[5]. Пізніше було виявлено, що бактеріальні клітини, в яких немає ядра, теж містять нуклеїнові кислоти.
Значення РНК в синтезі білків було припущено в 1939 році в роботі Торберна Оскара Касперсона, Жана Брачета і Джека Шульца.[6] Джерард Маірбакс виділив першу матричну РНК, що кодує гемоглобін кролика, і показав, що при її введені в ооцити утворюється той же самий білок.[7]
У Радянському Союзі в 1956—1957 роках проводилися роботи (А. Білозерський, О. Спірін, Е. Волкін, Ф. Астрахан) з визначення складу РНК клітин, які привели до висновку, що основну масу РНК в клітині становлять рибосомні РНК.
Северо Очоа отримав Нобелівську премію з медицини в 1959 році за відкриття механізму синтезу РНК.[8] Послідовність з 77 нуклеотидів однієї з тРНК дріжджів S. cerevisiae була визначена в 1965 році в лабораторії Роберта Холі, за що в 1968 році він отримав Нобелівську премію з медицини.[9] У 1967 Карл Воуз припустив, що РНК мають каталітичні властивості. Він висунув так звану Гіпотезу світу РНК, в якому РНК прото-організмів служила і як молекули зберігання інформації (зараз ця роль виконується ДНК) і молекули, яка каталізувала метаболічні реакції (зараз це роблять ферменти).[10]
У 1976 Волтер Фаерс і його група з Гентського університету (Бельгія) визначили першу послідовність геному РНК- яке міститься у вірусі, бактеріофага MS2. На початку 1990-х було виявлено, що введення чужорідних генів у геном рослин призводить до придушення вираження аналогічних генів рослини. Приблизно в цей же час було показано, що РНК довжиною близько 22 нуклеотидів, які зараз називаються мікро-РНК, відіграють регуляторну роль в онтогенезі нематод C. elegans. Гіпотеза про значення РНК в синтезі білків була висловлена Торбйорном Касперссоном (Torbjörn Caspersson) на основі досліджень 1937—1939 років.
Див. також
Примітки
- Сиволоб, 2008, с. 260.
- David L. Bentley (March 2014). Coupling mRNA processing with transcription in time and space. Nature reviews. Genetics 15 (3): 163–175. PMID 24514444. doi:10.1038/nrg3662.
- Сиволоб, 2008, с. 201-220.
- Takayuki Ohira & Tsutomu Suzuki (June 2011). Retrograde nuclear import of tRNA precursors is required for modified base biogenesis in yeast. Proceedings of the National Academy of Sciences of the United States of America 108 (26): 10502–10507. PMID 21670254. doi:10.1073/pnas.1105645108.
- Dahm R (2005). Friedrich Miescher and the discovery of DNA. Developmental Biology 278 (2): 274–88. PMID 15680349.
- Nierhaus KH, Wilson DN. Protein Synthesis and Ribosome Structure. — Wiley-VCH, 2004. — С. 3. — ISBN 3-527-30638-2.
- Carlier M (червень 2003). L’ADN, cette «simple» molécule. Esprit libre. Архів оригіналу за 23 серпня 2011. Процитовано ???.
- Ochoa S. (1959). Enzymatic synthesis of ribonucleic acid. Nobel Lecture. Архів оригіналу за 23 серпня 2011. Процитовано ???.
- Holley RW et al. Structure of a ribonucleic acid // Science. — 1965. — Т. 147, № 1664. — С. 1462–65. — DOI:.
- Szathmáry E. The origin of the genetic code: amino acids as cofactors in an RNA world // Trends Genet.. — 1999. — Т. 15, № 6. — С. 223–9. — DOI:.
Література
- Альбертс Б.; Брей Д.; Льюис Дж. и др. Молекулярная биология клетки в 3-х томах. — М.: Мир, 1994. — 1558 с. — ISBN 5-03-001986-3
- История биологии с начала XX века до наших дней. — М.: Наука, 1975. — 660 с.
- Льюин Б. Гены. — М.: Мир, 1987. — 544 с.
- Спирин А. С. Биосинтез белков, мир РНК и происхождение жизни
- Molecular Biology Of The Cell, 4е видання, 2002 р.
- А. В. Сиволоб (2008). Молекулярна біологія. К: Видавничо-поліграфічний центр "Київський університет".
Посилання
- Предсказание вторичной структуры РНК (англ.)
- База данных геномных тРНК (англ.)
- База данных псевдоузлов (англ.)
- Химики преодолели главное препятствие на пути к абиогенному синтезу РНК (рос.)
- Що таке ДНК та РНК — відео Tokar.ua