Нікотиновий ацетилхоліновий рецептор
Нікотиновий ацетилхоліновий рецептор (nACh-receptor) — підвид ацетилхолінових рецепторів, який забезпечує передачу нервового імпульсу через синапси і активується нікотином (крім ацетилхоліну). Цей рецептор входить до групи рецепторів-іонних каналів разом з ГАМКА-, гліциновим та серотоніновим 5-HT3- рецепторами.
Цей рецептор знайдено в хімічних синапсах як в центральній, так і в периферичній нервовій системі, в нервово-м'язових синапсах, а також в епітеліальних клітинах багатьох видів тварин.
Нікотиновий ацетилхоліновий рецептор було відкрито на початку 20-го століття як «рецепторну структуру нікотину», приблизно за 25-30 років до того, як була досліджена його роль у проведенні нервових сигналів, генерованих за допомогою ацетилхоліну. При потраплянні ацетилхоліну (ACh) на молекулу даного рецептору відкривається проникний для катіонів іонний канал, і це призводить до деполяризації клітинної мембрани та генерації нервового імпульсу в нейроні або скорочення м'язового волокна (у випадку нервово-м'язового синапсу).
Структура
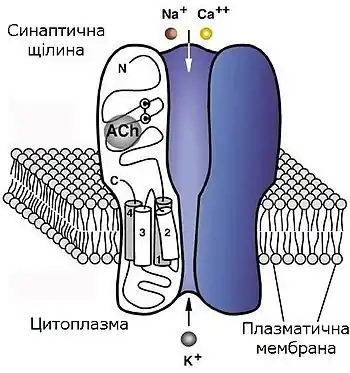
Висока концентрація нікотинових ацетилхолінових рецепторів в електричних органах деяких скатів, зокрема Torpedo californica та Torpedo marmorata, разом з виділенням α-бунгаротоксина (αBGT — поліпептида з яду змії Bungarus multicinctus, який є незворотнім лігандом та специфічним антагоністом ацетилхоліну для нікотинових рецепторів з нервово-м'язових синапсів), дозволила біохімічно очистити та детально дослідити нікотинові ацетилхолінові рецептори, а також ідентифікувати сайт зв'язування ацетилхоліну. Було показано, що даний рецептор є гетероолігомерним комплексом, який складається з чотирьох різних білкових субодиниць, котрі були названі відповідно до їхньої молекулярної маси (в кілодальтонах): α (40), β (50), γ (60), δ (65). При природній експресії в клітині спочатку виникають димерні комплекси α-γ та α-δ, потім формується тример α-β-δ, і врешті-решт, після поєднання димеру та тримеру, в клітинну мембрану вбудовується функціональний пентамер із стехіометрією α2βγδ.
Основний сайт зв'язування агоністу (ацетилхоліну) розташований на зовнішньо-клітинній поверхні кожної з α-субодиниць, поряд з сегментом М1, і оточений двома сусідніми залишками цистеїну (номери 192 та 193 в первинній структурі); для формування функціонального сайту зв'язування ці цистеїнові залишки мають бути поєднані дисульфідним містком між входячими до їхнього складу атомами сірки. Також для зв'язування ацетилхоліну важливим (але не завжди критичним) фактором є наявність в даному сайті залишків тирозину та триптофану. Сайт зв'язування ацетилхоліну сформований трьома паралельними α-спіралями білкової молекули, завдяки чому знаходиться в заглибленні між ними. Для відкриття іонного каналу рецептору необхідне зв'язування ацетилхоліну з обома сайтами, розташованими на α-субодиницях. Окремий зовнішньоклітинний домен на α-субодиниці, розташований в районі залишку Lys-125 на відстані близько 10 Å від сайту зв'язування ацетилхоліну, розпізнається не ацетилхоліном, а ендогенною сполукою 5-гідрокситриптаміном, а також особливим класом агоністів — похідних фізостигміну. Регіон поблизу залишку Lys-125 та сусідні частини рецепторної макромолекули, що включають дисульфідний місток (Cys128-Cys142), є дуже схожими у всіх субодиниць нікотинових рецепторів. З огляду на те, що β-, γ- та δ-субодиниці позбавлені агоніст-зв'язуючого сайту, вони називаються «структурними» субодиницями.
Трансмембранна частина рецептору утворює іонний канал, стінки якого сформовані сегментами М2 всіх п'яти субодиниць. Було доведено, що відносно невеликі пертурбації, а саме поворот на 4° двох агоніст-зв'язуючих субодиниць, призводять до значного зміщення сегментів М2 і відкриття пори іонного каналу, що є умовою виникнення катіонного струму через рецептор.
Фізіологія та фармакологія
Електрофізіологічна характеристика нікотинових рецепторів м'язової тканини вперше була дана завдяки внутрішньоклітинному відведенню електричних потенціалів; окрім того, нікотиновий рецептор був одним з перших, на яких вдалось записати електричні струми, що проходять через одиничний рецепторний канал. Використовуючи останній підхід вдалося показати, що іонний канал даного рецептору існує в дискретних відкритому та закритому станах. У відкритому стані рецептор може пропускати іони Na+ та К+ і, в меншій мірі, двохвалентні катіони; провідність іонного каналу при цьому є сталою величиною. Тим не менше, час існування каналу у відкритому стані є характеристикою, що залежить від напруги прикладеного до рецептору потенціалу, при цьому рецептор стабілізується у відкритому стані при переході від малих значень напруги (деполяризація мембрани) до великих (гіперполяризація). Довготривала аплікація ацетилхоліну та інших агоністів рецептора призводить до зниження його чутливості до рецепторної молекули і збільшення часу перебування іонного каналу в закритому стані — тобто у нікотинового рецептора спостерігається явище десенсетизації.
Класичною характеристикою нікотинових рецепторів в нервових гангліях та в головному мозку є холінергічні відповіді на електричне подразнення, які блокуються дигідро-β-еритроїдином; окрім того, для цих рецепторів характерне високоафінне зв'язування з тритій-міченим нікотином. αBGT-чутливі рецептори в нейронах гіпокампу характеризуються низькою чутливістю до ацетилхоліну, на відміну від αBGT-нечутливих рецепторів. Селективним та оборотним конкурентним антагоністом αBGT-чутливих рецепторів є метиллікаконітин, а деякі похідні анабезиїну завдають селективний активаційний вплив на цю групу рецепторів. Провідність іонного каналу αBGT-чутливих рецепторів є доволі високою (73pS); також їм притаманна відносно висока провідність іонів кальцію порівняно з іонами цезію (PCa/PCs 6.1 на фоні гіперполяризуючого утримуючого потенціалу). Цей рецептор виявляє незвичайні вольт-залежні властивості: загально-клітинний струм, записаний у фізіологічному стані, при накладенні деполяризаційних величин електричного потенціалу вказуює на достовірне зменшення проходження іонів через іонні канали; при цьому це явище регулюється концентрацією в розчині іонів Mg2+. Для порівняння, нікотинові рецептори на м'язових клітинах не виявляють ніякої зміни іонного струму при зміні значень мембранного електричного потенціалу, а NMDA-рецептор, котрому також притаманна висока відносна проникність для іонів Са2+ (PCa/PCs 10.1), виявляє зворотну картину зміни іонних струмів у відповідь на зміну електричного потенціалу та наявність іонів магнію: при підвищенні електричного потенціалу до гіперполяризуючих величин та підвищенні концентрації іонів Mg2+ іонний струм через даний рецептор блокується.
Інша важлива властивість αBGT-чутливих нейронних нікотинових рецепторів — це їхня реакція на стимуляцію. Експозиція високих концентрацій ацетилхоліну призводить до дуже швидкої десенсетизації відповіді окремого каналу та швидкого падіння електричної відповіді всієї клітини. Повторювальна експозиція коротких імпульсів ацетилхоліну також призводить до зменшення максимальної амплітуди рецепторної відповіді. При цьому енергетичне підживлення клітини високоенергомісткими молекулами (АТФ, фосфокреатин, креатин-фосфокіназа) або проміжними продуктами їхнього метаболізму здатне запобігти такому зменшенню. Судячи зі всього, зменшення загальної амплітуди клітинної відповіді віддзеркалює прогресуючий процес переходу все нових порцій рецепторів до деякого стабільного десенсетизованого стану, котрий може бути змінений на нормально функціонуючий тільки з витратами енергії. Майже всі аспекти функціонування αBGT-чутливих нікотинових рецепторів, включаючи ефективність агоністів, кооперативні ефекти, а також фракціонування за активністю та десенсетизація, регулюються зовнішньоклітинною концентрацією Са2+. Така регуляція може бути особливо важливою у випадках, коли рецептори розташовані на дендритах.
αBGT-нечутливі нейронні нікотинові рецептори мають високу спорідненість до ацетилхоліну, епібатидину та нікотину — риса, яка зумовлює їхню селективну активацію за допомогою екзогенних агоністів, що, в свою чергу, є причиною багатьох фізіологічних та поведінкових ефектів, зумовлених цим типом рецепторів. З огляду на те, що більшість (~90%) αBGT-нечутливих рецепторів в мозку мають будову α4β2, цей тип рецепторів досить яскраво виявляє багато ефектів, які спричинюються низькими концентраціями агоністу. З іншого боку, радіоактивно-мічений агоніст зв'язується з багатьма високоафінними сайтами в нервовій тканині, доводячи існування інших αBGT-нечутливих рецепторів. Велика кількість даних свідчить про те, що αBGT-нечутливі рецептори розташовані дискретно на кількох окремих типах пресинаптичних терміналів в специфічних ділянках мозку, де вони полегшують виділення частини нейромедіаторів. Порівняно з αBGT-чутливим рецептором, αBGT-нечутливий рецептор має нижчу проникність для Са2+, а його активація потенціюється при високих концентраціях Са2+. Більш того, αBGT-нечутливі нікотинові струми поділяються на чутливі та нечутливі до наявності в середовищі іонів Mg2+.

На додаток до селективної активації рецепторів ацетилхолін-подібними агоністами, всі підтипи нікотинових рецепторів активуються похідними фізостигміну; тим не менше, така активація притаманна тільки низькочастотним струмам одиничних рецепторів, котрі не можуть бути придушені антагоністами ацетилхоліну, а на загальноклітинному рівні не виявляється. Нещодавні дослідження доводять, що активність нікотинових рецепторів, отримана за допомогою ацетилхоліну або інших агоністів, може бути позитивно модульованою похідними фізостигміну, які зв'язуються із своїм специфічним сайтом на рецепторній молекулі. Досить можливою вважається наявність ендогенного ліганду даного сайту, і найвірогіднішим кандидатом на цю роль вважають 5-гірокситриптамін.
Нейроанатомія та патологія
Роль периферійних нікотинових рецепторів висвітлюється при аутоімунних захворюваннях, при яких вражаються окремі форми рецепторів. У пацієнтів, що хворіють на міастенію, антитіла до м'язових нікотинових рецепторів запобігають нормальному моторному функціонуванню м'язів, призводячи до загального послаблення (що є первинним симптомом даного захворювання).
Пухирчатка є дерматологічним ураженням, яке характеризується виникненням заповнених рідиною пухирів, що виникають завдяки порушенню клітинної адгезії. Нікотинові рецептори, яким притаманна композиція α3β2, відіграють важливу роль в процесі клітинної адгезії в кератиноцитах, і антитіла до цих рецепторів провокують виникнення пухирчатки.
В ЦНС дисфункція нікотинових ацетилхолінових рецепторів в гіпокампі та корі великих півкуль призводить до виникнення хвороби Альцгеймера.
Токсикологія
Токсини, які придушують функціонування нікотинових рецепторів, знайдені у багатьох представників рослинного та тваринного світу. Такий потужний ліганд, як анатоксин, що синтезується прісноводними синьо-зеленими водоростями, призводить до швидкої загибелі водних організмів при цвітінні води (тобто спалаху розмноження цих водоростей). Деякі рослинні токсини, такі як кураре та фізостигмін, що вражають нікотинові рецептори, використовуються людиною вже кілька тисячоліть. Метил-лікасонітин, що виробляється дельфінієм Delphinium elatum, є зворотнім конкурентним антагоністом з високою спорідненістю до αBGT-чутливих нейрональних нікотинових рецепторів. Відомо також, що нікотин сам по собі є речовиною, яка відповідає за виникнення тютюнової залежності. Дані щодо впливу на функції αBGT-чутливих рецепторів іонів Pb2+ можуть, як мінімум частково, пояснювати негативний вплив цього металу на пам'ять та вищу нервову діяльність мозку.
У відкритому стані нікотинові рецептори м'язового типу блокуються великою кількістю специфічних лігандів — блокаторів іонного каналу. До них належать місцеві анестетики (бупівакаїн, кокаїн, піперокаїн, лідокаїн), антимускаринові сполуки (атропін, скополамін), антагоністи з наркотичним ефектом (налтрексон), специфічні токсини (гістріонікотоксин), антивірусні речовини та антибіотики (амантадін, квінакрин), стимулятори (нікотин), психотропні речовини (кокаїн, РСР), та фосфор-вміщуючі органічні сполуки (диізопропілфлюорофосфат, зарин, VX). Незважаючи на різну хімічну природу, ці агенти-блокатори являють собою в більшості випадків молекули, позитивно заряджені на більшій частині поверхні. З огляду на те, що сайт, зв'язування з яким забезпечує блокування іонного каналу, є чутливим до заряду клітинної мембрани, дія блокаторів цього каналу є також електрозалежною; при гіперполяризаційних значеннях заряду мембрани блокування відбувається найактивніше.
Різновиди нікотинових рецепторів
Різновиди нікотинових рецепторів та їхні фармакологічні властивості
| Анатомічна локалізація | Субодиничний склад | ACh-чутливість (ЕС50) та кінетичні характеристики | Агоністи | Антагоністи |
| Електричні органи риб | α12β1γδ | 0.3μM | Карбамоілхолін | (+)-тубокукарин |
| М'язова система риб | α12β1εδ | - | Анатоксин | α-бунгаротоксин |
| ЦНС | α7 | 120μM, швидка десенситизація | Похідні анабазеїну | α-бунгаротоксин, метіллікоконітин |
| Автономні ганглії ЦНС | α4β2 | 0.3μM, повільна десенсетизація | Епібатідин, нікотин, n-етилкарбамоілхолін | дігідро-β-еритроідин, κ-бунгаротоксин |
| ЦНС | α3β4 | 0.1μM, повільна десенсетизація | Ацетилхолін | Мекаміламін |
| Кератиноцити | α3β2 | 0.1μM | Ацетилхолін, анатоксин, нікотин | Мекаміламін, κ-бунгаротоксин |
У людини відомо 9 альфа-субодиниць (CHRNA1, CHRNA2, CHRNA3, CHRNA4, CHRNA5, CHRNA6, CHRNA7, CHRNA9, CHRNA10), 4 бета-субодиниці (CHRNB1, CHRNB2, CHRNB3, CHRNB4), альфа2-дельта субодиниця, гамма- та епсилон-субодиниці.
Література
- Albuquerque EX, Pereira EFR, Castro NG, Alkondon M, Reinhardt S, Schröder G, Maelicke A (1995): Nicotinic receptor function in the mammalian central nervous system. Ann NY Acad Sci volume: 757 1995 pp 47-72
- Alkondon M, Albuquerque EX (1993): Diversity of nicotinic acetylcholine receptors in rat hippocampal neurons. I. Pharmacological and functional evidence for distinct structural subtypes. J Pharmacol Exp Ther volume: 265 pp 1455–1473
- Lena C, Changeux J-P (1993): Allosteric modulations of the nicotinic acetylcholine receptor. Trends Neurol Sci volume: 16 pp 181–186
- McGehee DS, Role LW (1995): Physiological diversity of nicotinic acetylcholine receptors expressed by vertebrate neurons. Annu Rev Physiol volume: 57 pp 521–546
- Pereira E, Reinhardt-Maelicke S, Schrattenholz A, Maelicke A, Albuquerque EX (1993): Identification and functional characterization of a new agonist site on nicotinic acetylcholine receptors of cultured hippocampal neurons. J Pharmacol Exp Therap volume: 265 1993 pp 1474–1491
- Schrattenholz A, Pereira EFR, Roth U, Weber K-H, Albuquerque EX, Maelicke A (1996): Agonist responses of neuronal nicotinic acetylcholine receptors are potentiated by a novel class of allosterically acting ligands. Mol Pharmacol volume: 49 pp 1-6
- Wonnacott S, Dasdo A, Sanderson E, Rowell P (1990): Presynaptic nicotinic receptors and the modulation of transmitter release. The Biology of Nicotine Dependence, Ciba Found Symp 152 pp 87-105 publisher: Wiley Chichester
