Адгезивні контакти
Адгезивні контакти (англ. adherens junctions, AJ) — якірні міжклітинні контакти асоційовані з мікрофіламентами, забезпечують цілісність та механічну міцність тканини, зокрема протистояння розтягу, та надають клітинам можливість координовано використовувати актиновий цитоскелет. Адгезивні контакти належать до гомофільних, тобто сполучають клітини однакового типу. В їх формуванні беруть участь білки кадгерини та катеніни[1][2].
Морфологічно адгезивні контакти є відносно простими, на відміну від десмосом, щільних та шілинних контактів, вони не мають високоспеціалізованих ультраструктур, окрім скупчення актинових філаментів. З поміж інших типів сполучень клітин вони вирізняються відносною гнучкістю та мінливістю[3].
Поширення
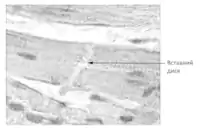
Найчастіше адгезивні контакти зустрічаються в епітеліальних тканинах, тут вони утворюють навколо кожної клітини поясок, який називають зоною прилипання (англ. zonula adherens). Такі зони в епітелії хребетних тварин переважно розміщуються базальніше від ділянки щільних контактів (англ. zonula occludens) і апікальніше від десмосом (англ. macula adherens)[1][2].
Проте поширення адгезивних контактів в організмі не обмежене тільки епітелієм, у багатьох неепітеліальних тканинах, наприклад у нервовій та сполучній, вони присутні у формі точкових або смужкоподібних сполучень клітин. Також вони добре виражені у серцевому м'язі, де забезпечують непряме сполучення скоротливого апарату кардіоміоцитів. Разом із десмосомами адгезивні контакти утворюють так звані вставні диски між клітинами міокарда[1][3].
Будова
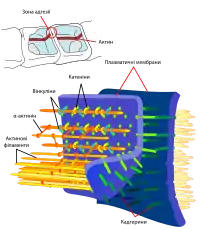
У зоні адгезивних контактів мембрани сусідніх клітин віддалені одна від одної на відстань 10—20 нм[3]. До складу адгезивних контактів входять три основні елементи:
- Власне сполучення клітин відбувається завдяки трансмембранним молекулам клітинної адгезії (англ. cell adhesion molecules, CAM), з яких найпоширенішими в адгезивних контактах є класичні кадгерини. Їхня N-кінцева позаклітинна частина у присутності іонів кальцію взаємодіє із подібною молекулою на сусідній клітині, таким чином забезпечуючи злипання клітин, а внутрішньоклітинний C-кінцевий домен зв'язується із якірними білками. Також в адгезивних контактах були виявлені інші трансмембранні білки: нектини та везатини[4].
- Адаптером між CAM та елементами цитоскелету виступають внутрішньоклітинні якірні білки, більшість з них кріплять цитоплазматичну сторону кадгеринів до актинових філаментів. Кадгерини безпосередньо взаємодіють із β-катенінами та плакоглобінами (γ-катенінами), які у свою чергу приєднуються до молекул α-катенінів, а ті — до вінкуліну, α-актиніну або ZO-1, які вже зв'язуються із актином[2][4]. У комплексі якірних білків з кадгеринами присутній також білок p120-катенін, що, ймовірно, бере участь у регулюванні сили адгезивних контактів між клітинами[1]. Нектини кріпляться до актинових філаментів через білок афадин[3][4].
- Третім компонентом є пучки актинових філаментів у сусідніх клітинах непрямо сполучені між собою. Також був продемонстрований зв'язок адгезивних контактів із мікротрубочками, хоча значного їх скупчення у цих ділянках і не спостерігається. Мікротрубочки можуть приєднуватись до адгезивних контактів як плюс-, так і мінус-кінцями, разом із моторними білками вони, ймовірно, беруть участь у транспортуванні білків, необхідних для формування контактів, зокрема кадгеринів[3].
Функції
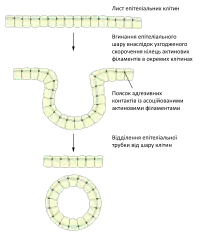
Однією із основних функцій адгезивних контактів є фізичне сполучення клітин у єдину тканину, їх послаблення часто приводить до дисоціації клітин. Такого ефекту можна досягнути обробивши тканину чи моношарову культуру хелатуючими агентами, такими як ЕДТА, що зв'язують іони кальцію, внаслідок чого взаємодія між кадгеринами порушується. Проте, хелатуючих агентів зазвичай недостатньо для повного розділення клітин, оскільки між ними існують інші — кальцій незалежні — контакти[3].
Адгезивні контакти забезпечують утворення широкої міжклітинної мережі із пучків скоротливих актинових філаментів, розташованих паралельно до мембран клітин і сполучених між собою за допомогою білків катенінів та кадгеринів. Така організація дозволяє не тільки протистояти механічному напруженню, а й узгоджувати поведінку клітин під час процесів морфогенезу. Наприклад, координоване скорочення кілець актинових філаментів сусідніх клітин є необхідним для формування трубок із листків епітелію, зокрема під час закладання нервової трубки[1]. Одним із прикладів є Shroom3-залежне скорочення зони прилипання, при цьому актин-зв'язуючий білок Shroom3 залучає до ділянки адгезивних контактів Rho кінази та активує міозин-II, внаслідок чого і відбувається скорочення[3].
Адгезивні контакти також задіяні у міжклітинній передачі сигналів, про це свідчить локалізація у зрілих контактах рецепторної тирозинфосфатази μ та білка RACK1, що взаємодіє з нею. При зниженні експресії α-катенінів спостерігається значене збільшення темпів проліферації епітеліальних клітин, було показано, що за цей ефект відповідає сигнальний шлях інсулін/MAPK[4].
Джерела
- Alberts B, Johnson A, Lewis J, Raff M, Roberts K, Walter P (2007). Molecular Biology of the Cell (вид. 5th). Garland Science. ISBN 978-0-8153-4105-5.
- Harvey Lodish et al. (2000). [http://www.ncbi.nlm.nih.gov/books/NBK21599/ 22.1 Cell-Cell Adhesion and Communication]. Molecular Cell Biology (вид. 4th). W H Freeman. ISBN 0-7167-3136-3. — Переглянуто 5 березня 2012
- Meng W, Takeichi M. (2009). Adherens junction: molecular architecture and regulation. Cold Spring Harb Perspect Biol. 6. PMID 20457565. doi:10.1101/cshperspect.a002899.
- Nagafuchi A. (2001). Molecular architecture of adherens junctions. Curr Opin Cell Biol. 13: 600–3. PMID 11544029. doi:10.1016/S0955-0674(00)00257-X.