Антитілозалежне посилення інфекції
Антитілозалежне посилення інфекції (англ. antibody-dependent enhancement, ADE) — явище, при якому зв'язування вірусу з субоптимальними нейтралізувальними або не нейтралізувальними антитілами викликає його проникнення в імунні клітини інфікованого організму і вірусну реплікацію[1][2]. ADE може проявлятися в процесі розвитку первинної або вторинної вірусної інфекції, а також після вакцинації при подальшій інфекції[3][4][5].
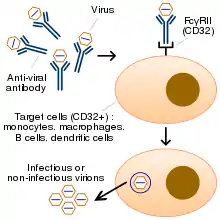
Антитіла, за своєю природою, сприяють фагоцитуванню вірусу імунними клітинами. Зазвичай фагоцитування комплексу вірус-антитіло супроводжується деградацією вірусу, але може, в разі антитілозалежного посилення інфекції, викликати, навпаки, реплікацію вірусу, з подальшою загибеллю імунних клітин. Таким чином, вірус як би «обманює» процес фагоцитозу імунних клітин і використовує антитіла господаря в якості «троянського коня». Механізм антитілозалежного посилення інфекції, який включає фагоцитоз імунних комплексів через рецептор FcγRII / CD32, вивчений краще, ніж механізм, що включає фагоцитування через рецептор комплементу. Клітини, що експресують FcγRII/CD32 рецептор, представлені моноцитами, макрофагами, деякими категоріями дендритних клітин, B-клітинами та іншими. Антитілозалежне посилення інфекції ускладнює розробку противірусних вакцин.
Так, при зустрічі з інфекцією деякі вакцини, через механізм антитілозалежного посилення інфекції, замість захисту здатні провокувати важчий перебіг захворювання.
Історія
Феномен ADE вперше описав Ройл Хоукіс (R. A. Hawkes[6]) у 1964 році: він виявив підвищення продукції різних флавивірусів у клітинах курячого ембріона, вперше експонованих до вірусів, що перебувають в середовищі з низьким вмістом специфічних антитіл[7]. Згодом він навів докази, що збільшення «виходу» вірусу в подібних експериментах викликане утворенням комплексу «вірус-антитіло»[8].
Проте, наприкінці 1960-х і початку 1970-х років вже іншими дослідниками виявлена роль ADE в патогенезі важких форм геморагічної лихоманки, викликаної вірусом лихоманки денге. Було встановлено, що наявність антитіл в сироватці крові видужалого, що залишилися після легко перенесених випадків лихоманки денге, призводить до важкого перебігу хвороби, якщо відбулося повторне зараження, але вірусом лихоманки денге іншого серотипу[9][10].
Ця проблема призводить до того, що створити безпечну вакцину проти вірусу лихоманки денге вкрай складно[11]. Компанія Санофі Пастер (Франція) спробувала це зробити і створила вакцину, яка містила антигени до всіх чотирьох серотипів вірусу лихоманки денге. Вакцина цієї компанії отримала назву Денгваксія (Dengvaxia)[12]. Серйозним недоліком Денгваксії є те, що вона діє точно так само, як природна первинна інфекція, і різко збільшує ризик розвитку важкої вторинної інфекції, яка пов'язана з важчою формою лихоманки денге. Причина цього явища, на думку авторів огляду літератури, пов'язана з явищем антитілозалежного посилення інфекції.
Так, в рамках клінічного дослідження була проведена вакцинація понад 10 000 дітей у віці 2-14 років в п'яти країнах Азіатсько-Тихоокеанського регіону. Аналіз даних, який провели фахівці з компанії Санофі Пастер, показав, що вакцинування дітей, яким було менше ніж 9 років, призводило до їхнього частого важкого захворювання і госпіталізації через лихоманку денге. Причому частота цієї госпіталізації в два рази перевищувала частоту госпіталізації дітей з контрольної групи, в якій діти не були вакциновані[12]. В іншій статті[13] дані пояснювалися тим, що чим молодша дитина, тим менш імовірно, що вона вже стикалася з інфекцією денге і тим ймовірніше, що вакцина зробила її зіткнення з інфекцією набагато небезпечнішим в порівнянні з невакцинованою дитиною.
В науковому журналі «New England Journal of Medicine» повідомляється, що департамент охорони здоров'я Філіппін розпочав програму масової вакцинації Денгваксією у квітні 2016 року, яка охопила приблизно 830 тисяч дітей, але призупинив програму наприкінці 2017 року. Зупинка програми вакцинації сталася, коли Санофі Пастер оприлюднила результати вакцинації. Виявилося, що вакцина певною мірою захищала дітей старшого віку, але часто шкодила здоров'ю дітей молодшого віку, у яких не було імунітету до вірусу лихоманки денге[14]. Згідно з рекомендаціями ВОЗА існує підвищений ризик госпіталізації та розвитку важкої форми лихоманки денге у серонегативних осіб, починаючи приблизно через 30 місяців після отримання першої дози вакцини[15]. Серонегативними називаються люди, які не мають антитіл на певний інфекційний агент. Історія набула широкого міжнародного розголосу як "Dengvaxia controversy«(також "Dengvaxia issue" або " Dengvaxia mess»)[16][17][18].
Наразі вакцина Денгваксія, згідно з інформацією Центру по запобіганню інфекційних захворювань США (CDC), рекомендується до застосування тільки людям, серопозитивним щодо вірусу лихоманки денге, тобто людям, які вже стикалися з відповідним вірусом і виробили антитіла[19]. Вважається, що у цих людей вакцина запобіжить захворюванню при зустрічі з новою інфекцією або послабить тяжкість його перебігу. Однак пошуки безпечної та ефективної вакцини проти вірусу денге тривають і, не виключено, що вони увінчаються успіхом[20][21].
«Список вірусів людини та тварин, які можуть викликати ADE, досить довгий і включає віруси грипу А, віруси Коксакі, респіраторно-синцитіальний вірус, вірус Ебола та ін.»[22][23][24].
З початку 2020 року, коли в світі стала стрімко поширюватися епідемія COVID-19[25], викликана коронавірусом SARS-CoV-2, з 11 березня 2020 року охарактеризована Всесвітньою організацією охорони здоров'я (ВООЗ) як пандемія COVID-19[26][27], інтерес до феномену антитілозалежного посилення інфекції (ADE) багаторазово зріс. Це пов'язано, в першу чергу, із заявленим початком розробки різними провідними науковими центрами вакцини проти нового коронавірусу, де одним з найважливіших факторів, що визначають її безпеку, є відсутність феномену ADE[28]. Обговорення проблеми антитілозалежного посилення інфекції при розробці вакцин проти SARS-CoV-2 і терапії моноклональними антитілами детально викладено в опублікованій роботі в журналі Nature[29]. Так само ця проблема піднімається в іншій статті в журналі Nature Biotechnology[30].
Дискусія щодо еффектів послаблення імунітету та ADE в контексті пандемії Covid-19 почалася навесні 2021, з посиленням епідемічної обстановки у країнах, які використовували вакцину Sinovaс. Зокрема, після вакцинації фіксувалося збільшення кількості підтверджених випадків та поширенні коронавірусу в Чилі, Туреччині та Пакистані. У лютому Чилі запровадила близько 9 млн доз вакцини Sinovac, у середньому 47 доз на 100 осіб, це найвищий рівень вакцинації у Південній Америці. Однак кількість позитивних результатів, що були в Чилі, зросла, а не впала. Туреччина почала вводити вакцину Sinovac у середині січня, вакцинували не менше 8 млн осіб (понад 10 % населення). Але кількість хворих відновилася наприкінці лютого, а 30 березня було зареєстровано 37303 нових випадків, найвищий показник за один день. У Пакистані, який використовував вакцину з початку лютого, національний коефіцієнт позитивності тестів підвищився до 11 % (найвищий рівень з моменту спалаху)[31]. 31 жовтня 2021 року семиденний середній показник поширення коронавірусу в Німеччині також підвищився і склав 145,1 випадка на 100 тис.жителів. Всього за той же період RKI зафіксував більше 2,2 млн нових випадків зараження. Тобто доля повністю вакцинованих від числа всіх інфікованих склала 5 %.[32]. Таким чином, виникло особливе занепокоєння ймовірністю того, що вакцина може не захищати від коронавірусу, а навпаки, посилювати його дію. Однак досі невідомо першопричину даних явищ, серед гіпотез виокремлюють гіпотезу еффектів послаблення імунітету та гіпотезу ADE. У детальному огляді випадків ADE, потенційно пов'язаних з вакцинами проти SARS-CoV-2, автори зазначили: «В даний час відсутні відомі клінічні дані, імунологічні аналізи або біомаркери, що дозволяють диференціювати важку вірусну інфекцію, викликану захворюванням з посиленим імунітетом шляхом вимірювання антитіл, Т-лімфоцитів або внутрішньої відповіді господаря»[29].
Антитілозалежне посилення інфекції у коронавірусів
Феномен антитілозалежного посилення інфекції описаний для вірусів з геном, несучим (+) ланцюг РНК, включати коронавіруси[33][34][35][36][37]. У коронавірусів це явище викликають антитіла до шиповидного (S) білку. Не виключено, що антигенна мінливість S-білка, яку можуть викликати різні варіанти глікозилювання та / або зміни конформацій, сприяє ADE.
Антитілозалежне посилення інфекції у альфа-коронавірусів
Вірус інфекційного перитоніту кішок (Feline infectious peritonitis (FIP) або (FIPV))[38][39] являє собою альфа-коронавірус, яким часто заражаються як домашні, так і дикі кішки[40]. У багатьох тварин захворювання проходить безсимптомно або викликає слабкий кишковий розлад. Однак у деяких кішок розвивається перитоніт, який майже завжди призводить до летального результату. Вірус FIPV існує в двох формах, які називають біотипами або патотипами. Один з біотипів відповідає за безсимптомне захворювання, а інший за важке. Передбачається, що два біотипи розрізняються генетично, проте довести цю гіпотезу поки не вдалося[41]. Тому в реальності остаточно не відомо, чим відрізняються біотипи. Вакцинація проти FIPV може посилювати тяжкість захворювання і призводити до летального результату[42]. Цю проблему пояснюють ADE, оскільки показано, що in vitro інфікування макрофагів вірусом FIPV може бути ініційовано моноклональними антитілами, націленими на шиповидний S-білок[43][44]. Цікаво, що в основному ADE викликали антитіла підкласу IgG2а, здатні нейтралізувати вірус, в той час як протестовані антитіла підкласу IgG1 не викликали такого ефекту. Вірусне зараження макрофагів і моноцитів in vitro спостерігали і з антитілами з сироваток кішок, інфікованих вірусом[45]. Ефект ADE також пояснює, чому у половини кішок після експериментального зараження FIPV розвивається перитоніт у разі, якщо їх попередньо пасивно імунізували антивірусними антитілами[46]. У деяких країнах виробляється вірусна вакцина проти FIPV, заснована на аттенуйованому вірусі і застосовується у формі крапель для носа. Однак використання цієї вакцини, як з точки зору безпеки, так і ефективності, залишається спірним питанням[47]. Цікаво, що вакцинація векторним конструктом, що експресує N-білок вірусу, призводить до кращих результатів: при зараженні вірусом вакцинованих велика частина кішок залишається живими[48].
Антитілозалежне посилення інфекції у бета-коронавірусів
Деякі бета-коронавіруси здатні провокувати антитілозалежне посилення інфекції. Це явище для вірусів SARS-CoV-1 і MERS-CoV, що викликають відповідно важкий гострий респіраторний синдром (SARS) і Близькосхідний респіраторний синдром (MERS), описано в безлічі опублікованих робіт[49][50][51][52][34]. Явище ADE було продемонстровано як у клітинних культурах, так і у модельних тварин, але не у людини. Вважається, що S-білок вірусу безпосередньо пов'язаний з цим явищем. Антитіла, вироблені на вакцинний варіант вірусу з одними антигенними детермінантами S-білка, можуть втратити властивість нейтралізувати вірус при інфекції вірусами з видозміненим білком. Такі антитіла можуть як і раніше зв'язуватися з вірусом, але при цьому мати меншу афінність і утворювати менш стабільні комплекси в порівнянні з комплексами, які вони утворюють з «вакцинною» формою вірусу. В результаті комплекс антитіло-вірус може виступати в якості «троянського коня», допомагаючи вірусу проникнути в моноцити, макрофаги (а також інші імунні клітини господаря), запускаючи в цих клітинах інфекційний процес. Можливо, існує причинно-наслідковий зв'язок між титром IgG антитіл до епітопів шиповидного S-білка і системним запаленням у дітей, які заразилися вірусом вдруге. Наприклад, на моделях приматів було показано, що IgG антитіла до пептиду s597–603 S-білка для вірусу SARS-CoV-1 викликають виробництво антитіл, здатних провокувати ADE.[53]
Інфекція імунних клітин in vitro
Так, антитіла до S-білка SARS-CoV-1 сприяють проникненню вірусу у В-клітини[37][54], моноцити[50][51] і макрофаги[49]. У цих клітинах вірус реплікується, але не дає продуктивної інфекції. Це може бути пов'язано з тим, що заражені імунні клітини не експресують в достатній кількості серинових протеаз, необхідних для активації віріонів. Однак, не виключено, що неактивні віріони можуть активуватися і ставати інфекційними при проникненні в клітини дихального епітелію, в мембранах яких присутні потрібні для активації протеази. У той же час, реплікація вірусу, навіть без утворення інфекційних віріонів, може призводити до масової загибелі імунних клітин, що несуть рецептор Fc𝛾RIIγ. Деякі моноклональні антитіла до S-білка SARS-CoV-1[55] і МERS-CoV[56] також провокують ADE.
Імунопатологія у модельних тварин після вакцинації
Існують приклади того, що антитіла класу IgG на антигени S-білка SARS-CoV-1 викликають важке пошкодження легенів, опосередковане макрофагами у макак[57]. Незважаючи на те, що вакцинація векторним конструктом знижувала вірусне навантаження після зараження SARS-CoV-1, наявність IgG антитіл до S-білка у імунізованих макак значно посилювало запальне пошкодження легенів при реальній інфекції. Цікаво, що вірусна інфекція після вакцинації антигенами S-білка інших модельних тварин призводила до подібних негативних результатів. Наприклад, у тхорів при зустрічі з реальною вірусною інфекцією SARS-CoV-1 після вакцинації рекомбінантною вісповакциною, що експресує S-білок, виникав важкий гепатит[58]. У мишей виникало важке легеневе запалення знову ж при зустрічі з вірусною інфекцією після вакцинації інактивованим вірусом[59] або чотирма іншими варіантами вакцинного матеріалу[60]. Вакцинація мишей векторним конструктами, що експресують N-білок вірусу SARS-CoV-1 також викликала імунопатологію при зараженні тварин інфекційним вірусом[61][62].
ADE може виникнути під час первинної інфекції або при повторному зараженні після природної інфекції. У кроликів, інтраназально інфікованих MERS-CоV, розвивалася легенева патологія, що характеризується віремією і важким запалення легенів. При повторному зараженні MERS-CоV, незважаючи на наявність антитіл, кролики хворіли знову і пошкодження легенів були більш важкими, ніж під час первинної інфекції[63]. Інфекція вірусами SARS-CоV-1[60] або MERS-CoV[64] викликала більш важку пневмонію у вакцинованих тварин, незважаючи на високий рівень специфічних нейтралізуючих антитіл. У людей иммунодомінантний епітоп SARS-CoV-1 S-білка індукував продукцію як специфічних нейтралізуючих антитіл, так і антитіл, що підсилюють інфекцію макрофагів in vitro[65].
Механізм антитілозалежного посилення інфекції у коронавірусів
Існують різні гіпотези про те, як відбувається ADE, і цілком імовірно, що існує більше одного механізму. Нижче описаний механізм, який пов'язаний з FcγRII рецепторами імунних клітин і S-білком коронавірусів.
Рецептор імунних клітин FcγRII / СD32 сприяє їх зараженню і ADE
Було показано, що специфічні антитіла (IgG) при ADE формують недосконалі, неміцні комплекси з вірусом, допомагаючи йому заражати імунні клітини господаря, що несуть рецептор Fc𝛾RII[35][50][51]. Клітини, що експресують цей рецептор (FcγRII/СD32), представлені моноцитами, макрофагами, деякими категоріями дендритних клітин і B-лімфоцитів. Комплекс антитіла з вірусом зв'язується з Fc𝛾RII рецептором і фагоцитується СD32 + клітинами[52][34][56]. У нормі цей процес призводить до руйнування вірусу всередині імунної клітини і одужання. Однак при патології, вірус, звільнившись від антитіла, починає реплікативний цикл всередині імунної клітини, що поглинула його[49]. Цей процес описаний для різних альфа-і бета-коронавірусів, включаючи SARS-CoV-1. Він може призводити до масової загибелі імунних клітин і, як наслідок цієї загибелі, викликати цитокіновий шторм.
Специфічні нейтралізуючі антитіла зв'язують вірус набагато міцніше, і вірус повністю втрачає здатність інфікувати клітини. Більш того, вірус, будучи всередині моноциту або макрофага, не може вивільнитися після поглинання комплексу вірус-антитіло і піддається руйнуванню. Таким чином, комплекс вірусу з специфічними нейтралізуючими антитілами призводить до елімінації вірусу з організму, а комплекс з недосконалими антитілами, у яких константа зв'язування (константа асоціації, Ка) нижче в порівнянні з нейтралізуючими антитілами, — до реплікації вірусу в клітинах імунної системи, посилення інфекції і можливого цитокінового шторму.
Експресія двох видів рецепторів FcγRIIa і FcγRIIb, але не FcγRI або FcγRIIIa, індукувала ADE, викликане SARS-CoV-1[66]. При цьому було показано, що тяжкість захворювання SARS залежить від алельного поліморфізма FcγRIIa; у індивідуумів з ізоформою Fcγriia рецептора, який взаємодіє як з IgG1, так і з IgG2, розвивається більш важке захворювання, ніж у індивідуумів з ізоформою Fcγriia рецептора, який зв'язується тільки з IgG2[67].
IgG антитіла відповідають за ADE
FcγRII рецептори зв'язують тільки IgG антитіла[68]. У деяких експериментах було показано, що ADE в основному викликається антитілами підкласу IgG2a, в той час як тестовані антитіла підкласу IgG1 не викликали такого ефекту[69].
Універсальний механізм ADE, опосередкований IgG антитілами до S-білка
На підставі аналізу літератури[70] деякі дослідники роблять висновок про те, що для SARS-CoV-1, MERS-CoV існує універсальний механізм зараження імунних клітин, який призводить до ADE. Тільки антитіла, націлені на S-білок, але не на інші вірусні білки, здатні формувати комплекси з коронавірусом, які фагоцитуються імунними клітинами і провокують вірусну реплікацію, замість вірусного руйнування[49][34][56][65]. По всій видимості, S-білок відрізняється від інших вірусних білків тим, що він здатний змінювати імунодомінантні антигенні детермінанти за рахунок зміни конформації рецептор-зв'язуючого домену (RBD) і типів глікозилювання.
У роботі показано[56], що моноклональні нейтралізуючі антитіла, специфічні до RBD, опосередковують проникнення вірусу MERS-CoV в імунні клітини, функціонально імітуючи вірус-специфічні рецептори. Автори вважають, що антитіла, спрямовані проти інших ділянок S-білка і не пов'язані з його конформаційними змінами, з меншою ймовірністю призводитимуть до ADE. Також показаний доза-залежний ефект ступеня тяжкості ADE від концентрації антитіл.
Структура і конформаційна мінливість S-білка
S-білок бетакоронавірусів існує у вигляді тримера і складається з трьох ланцюгів, кожен з яких, в свою чергу, утворює дві субодиниці S1 і S2[71]. Субодиниця S1 несе рецептор-зв'язуючий домен (RBD). Між субодиницями S1 і S2 знаходиться сайт розрізання сериновими протеазами.[72] Вірус набуває здатність інфікувати клітини тільки після того, як відбувається протеолітичне розщеплення і кожна молекула білка розділяється на дві субодиниці. S1 може перебувати в двох конформаціях — відкритій і закритій. Біофізичне дослідження структури S-білка SARS-CoV-2 і аналіз структури з роздільною здатністю в 3.5A показали, що найбільш часто зустрічається S-білок, у якого одна з молекул тримера RBD знаходиться у відкритій конформації.
Антитіла або клітинний імунітет до N-білків коронавірусів можуть провокувати пневмонію
Вакцинація векторним конструктом, що експресує N-білок SARS-CoV-1, сприяє розвитку важкої пневмонії у мишей, після інфекції SARS-CoV-1[62]. Можливо, ця пневмонія пов'язана з ADE.
N-білок може посилювати пневмонію
Хоча можливий і інший механізм, так було показано, що n-білки SARS-CoV, MERS-CoV і SARS-CoV-2 можуть зв'язуватися з сериновою протеазою MASP-2, яка бере участь в шляху активації комплементу. Зв'язування викликає індуковану білком гіперактивацію комплементу. Гіперактивація посилює індуковану запальну пневмонію у мишей, тому можна припустити, що воно буде викликати аналогічні проблеми у людей. Мотив N-білка (115—123) безпосередньо взаємодіє з MASP-2[73].
Можлива роль ADE в патогенезі SARS і COVID-19
Патогенез захворювання SARS і COVID-19, на думку авторів деяких робіт[74][75][76][77][78][79][80], пов'язаний з ADE, що виявляється в інфекції макрофагів, В-клітин і моноцитів. Автори робіт вважають, що ця інфекція є ключовим кроком у розвитку хвороби та її еволюції від легкої форми до важкої з критичними симптомами. ADE може пояснити спостережуване порушення регуляції імунітету, включаючи апоптоз імунних клітин, що сприяє розвитку Т-клітинної лімфопенії (лімфоцитопенії)[81], запальний каскад з накопиченням макрофагів і нейтрофілів в легенях, а також цитокіновий шторм[70]. Раніше інші дослідники теж висловлювали схожу гіпотезу щодо SARS[37][82].
Вен Ші Лі та інші вказали, що тяжкість захворювання та імунна відповідь пов'язані з вірусним навантаженням нелінійно: «пацієнти з симптомами показали більш високі титри антитіл до SARS-CoV-2 і швидше виводили вірус з верхніх дихальних шляхів». Вони також відзначили, що дані досліджень свідчать: Т-клітинні відповіді проти SARS-CoV-2 можуть бути виявлені на високому рівні при легких і безсимптомних інфекціях, тоді як сильні титри антитіл більш тісно пов'язані з важким COVID-19". Вони підкреслили важливість збору інформації про механізми ADE при COVID-19 для забезпечення безпеки вакцин при їх широкому застосуванні[79].
Динаміка виробництва IgG антитіл, націлених на S-білок у хворих SARS
На користь того, що антитіла до S-білка можуть шкодити пацієнтам, викликаючи ADE, говорять і наступні спостереження, зроблені на невеликій групі пацієнтів з шести осіб, три з яких одужали, а три померли. Порівняльний аналіз специфічної гуморальної відповіді показав, що у пацієнтів, які померли від SARS-CoV-1 інфекції, нейтралізуючі антитіла до S-білка вироблялися значно швидше, ніж у видужалих людей[83]. Так було виявлено, що на 15-й день захворювання у пацієнтів, згодом померлих, титр антитіл до S-білка був значимо вище, ніж у тих, хто згодом одужав. При цьому, хоча титр нейтралізуючих антитіл протягом захворювання у згодом померлих пацієнтів зростав швидше в порівнянні з титром у згодом видужали пацієнтів, він також швидше падав. У той же час, у пацієнтів, які згодом одужали, титр антитіл збільшувався повільніше, але виростав до більш високого рівня і довше тримався на цьому рівні. Така динаміка зміни титрів антитіл була характерна як для IgM, так і IgG антитіл. Можна припустити, що у пацієнтів, згодом померлих, розвинулося антитілозалежне посилення вірусної інфекції у важкій формі і швидке вироблення антитіл до S-білка, які не могли нейтралізувати вірус, сприяло цьому. Можливо, уповільнене зростання титру сприяло виробленню антитіл з більш високою константою зв'язування, що відповідає міцнішим комплексам антиген-антитіло, з афінністю і авідністю, достатніми для нейтралізації вірусу. Значуще перевищення рівня антитіл у важких хворих порівняно з не важкими спостерігалося і на вибірці з 325 пацієнтів в іншій роботі[84]. Такі ж дані отримали й інші дослідники на вибірці з 347 хворих SARS. Більш того, було виявлено, що у пацієнтів, згодом померлих, антитіла з'являлися раніше за все[85].
Мінливість S-білка може забезпечувати антигенну різноманітність, що провокує ADE
Вірус зі зміненими антигенними властивостями S-білка може взаємодіяти з нейтралізуючими антитілами, виробленими на початковий варіант вірусу, утворюючи менш стабільні комплекси. Ці комплекси можуть «затягувати» вірус в моноцити, макрофаги або В-клітини, де той, вивільняючись з комлекса, може реплікуватися. При цьому може розвиватися генералізована інфекція (i. generalisata) і цитокіновий шторм[86]. Не виключено, що вірус змінює антигенні детермінанти за рахунок мутацій[87], зміни глікозилювання[88] та/або конформації S-білка[71].
Антитілозалежне посилення інфекції при ВІЛ-інфекції
Першими на антигенний імпринтинг при розробці ВІЛ- вакцин ще на початку 1990-х рр. у буквальному сенсі «натрапили» P.L. Nara et al.[89] Про існування даного феномена вони не підозрювали. Їх метою було розширення імунної відповіді на антигени ВІЛ щодо вірусів близьких серотипів різного географічного походження. Ввівши шимпанзе глікопротеїд gp120, отриманий з штаму ВІЛ-1 IIIB, і провівши через 175 діб. повторну вакцинацію gp120, виділеним з штаму ВІЛ-1 RF, що має інше географічне походження, дослідники несподівано для себе виявили зростання титрів антитіл до gp120 штаму IIIB і відсутність захисного ефекту при зараженні тварин ВІЛ-1 RF. Проведений ними ретроспективний аналіз наукової літератури показав, що феномен антигенного імпринтингу вже був описаний для інших ретровірусних інфекцій, зокрема, що викликаються вірусом вісни у овець[90] і вірусом інфекційної анемії у коней[91].
При клінічному вивченні протективного ефекту ВІЛ-вакцини, що включає в якості антигенного компонента gp120.16, виділений з ВІЛ-1 SF2, отримані подібні результати. Люди, вакциновані такою вакциною і які мають високі титри антитіл до gp120.16, виявилися сприйнятливі до варіантів ВІЛ-1, що циркулюють в їх популяції. При розвитку у вакцинованих ВІЛ-інфекції, в сироватці їх крові переважали антитіла до gp120.16 ВІЛ-1 SF2, а не до такого ж оболонкового глікопротеїну вірусу, що викликав інфекцію[92].
N. Larke et al.[93] у дослідах на мишах виявили, що включення в Експериментальні ВІЛ-вакцини антигенних білків ВІЛ різних клад (clade), «глушить» індукцію Т-клітинних відповідей на інші епітопні варіанти антигенів вірусу. Феномен антигенного імпринтингу виявлений і при вивченні імунної відповіді у ВІЛ-інфікованих пацієнтів. Вироблення антитіл на ВІЛ у них має олігоклональний характер. Одночасно відбувається порушення співвідношення κ/λ типів легких ланцюгів антитіл, що підтримується протягом багатьох років незалежно від швидкості прогресування захворювання. Обмежені (restricted) і при цьому стабільно підтримувані антитільні відповіді на антигени ВІЛ у таких пацієнтів являють собою одну з причин неможливості вироблення плазмоцитами антитіл до ВІЛ-1, які б ефективно пов'язували сероваріанти вірусу, що утворилися в ході персистуючого інфекційного процесу[94].
Антитілозалежне посилення інфекції при малярії
Розробка безпечної вакцини від малярія, що ефективно блокує вторгнення малярійного плазмодія (Plasmodium falciparum) в еритроцити людини є дуже важливим завданням біотехнології[95][96]. Особливо така вакцина потрібна для людей які тривалий час живуть в ендемічному по малярії регіоні. Однак на шляху створення безпечної та ефективної вакцини є серйозні перешкоди в тому числі у вигляді антигенного імпринтингу який приводив у модельних тварин до антитіло-залежного посилення інфекції, викликаного вакцинацією.
У малярійного плазмодія є безстатеві клітинні форми, які отримали назву мерозоїти. Зараження людини починається з укусу комара-переносника. У момент і після укусу спорозоїти, які представляють із себе певну стадію розвитку малярійного плазмодія, з слинних залоз комара проникають в кров людини і з потоком крові добираються до печінки, де впроваджуються в гепатоцити. Протягом наступного періоду часу в гепатоцитах відбувається утворення мерозоїтів, що представляють із себе іншу клітинну стадію розвитку малярійного плазмодія. Мерозоїти проникають в еритроцити і починають розмножуватися безстатевим шляхом. При розриві еритроцитів мерозоїти потрапляють в кров, що призводить до періодичних нападів лихоманки.
Те що перехресні імунні реакції на збудники малярії можуть спотворити репертуар Т- і В-клітин, індукованих малярійними спорозоитами і мерозоїтами, і вплинути на індукцію захисного імунітету у відповідністтю з концепцією антигенного імпринтингу («первинного антигенного гріха»), тобто відповісти на варіант плазмодія, який викликав інфекцію раніше, а не на той, який присутній в організмі і розмножується в даний момент, відомо з початку 1990-х рр.[97].
Антигенному імпринтингу при малярії сприяє антигенна різноманітність поверхневих білків у спорозоїтів і мерозоїтів — наслідок множинних точкових мутацій або варіацій числа, довжини і послідовності амінокислотних повторів. Ці білки процесуються і презентуються антигенпрезентуючими клітинами людини і активують Т- і В-клітини. Окремі клони Т- і В-клітин можуть стати домінуючими в імунній відповіді на наступні малярійні інфекції, але не обов'язково ця відповідь буде захисною[98].
Саме поверхневі білки мерозоїтів є важливою мішенню для розробки вакцини. Завдяки роботам R. J. Pleass et al. (2003),[99] вдалося показати можливість створення протималярійної вакцини на основі 19-кДа фрагмента білка MSP119, що знаходиться на поверхні мерозоїтів. Зв'язування специфічних антитіл з білком MSP119 може блокувати проникнення збудника малярії в еритроцити, активуючи його знищення фагоцитами людини. Однак такий сценарій реалізується не завжди. Вакцинація білком MSP119 іноді запобігає захворюванню в популяції людей, а іноді вона його підсилює. Досліди на мишачих моделях[100] допомогли розібратися в проблемі. Дослідники на тваринах моделювали відповідь на вакцинацію рекомбінантним білком MSP119. Моделювання несподівано показало, що зараження еритроцитів мишей малярійним плазмодієм може сприяти утворенню антитіл до рекомбінантного білка MSP119, який містила вакцина. Титр антитіл, після перенесеної мишами експериментальної малярії, можна було підвищити вакцинацією рекомбінантним білком MSP119. Однак дія, виконана у зворотному порядку, тобто спочатку одноразова ін'єкція рекомбінантного білка MSP119 (субоптимальна вакцинація), а потім інфікування малярійним плазмодієм призвела до утворення антитіл до MSP119, що не володіють протективною дією. Навпаки, ці антитіла сприяли зараженню мишей збудником малярії, так як мали змінену специфічність. Антитіла до MSP119 утворилися, але вони не володіли протективною дією, і приводили до зниження природного імунітету до зараження збудником малярії. Схожі результати були отримані і в іншій роботі. Так було показано, що антитіла проти поверхневих білків мерозоїту посилюють його проникнення в еритроцити in vitro і in vivo[101]. Це відбувається завдяки зв'язуванню антитіл з рецептором комплемента 1 (СR1), що говорить про тісний зв'язок між феноменами антигенного імпринтингу і антитілозалежного посилення інфекції. Проблеми, пов'язані з розробкою протималярійної вакцини детально описані в огляді літератури[102].
Труднощі непрогнозованості імунних реакцій як у осіб, що живуть в ендемічних по малярії регіонах, так і тих, що потрапили туди без перенесеної малярії в минулому, залишилися навіть після встановлення ролі білка MSP119 в імунній відповіді.
Примітки
- Sol M. Cancel Tirado, Kyoung-Jin Yoon. Antibody-Dependent Enhancement of Virus Infection and Disease // Viral Immunology. — 2003. — Т. 16, вип. 1 (1 квітня). — С. 69–86. — ISSN 0882-8245. — DOI:.
- Matthew Zirui Tay, Kevin Wiehe, Justin Pollara. Antibody-Dependent Cellular Phagocytosis in Antiviral Immune Responses // Frontiers in Immunology. — 2019. — Т. 10 (15 лютого). — ISSN 1664-3224. — DOI:.
- Khandia, R.; Munjal, A.; Dhama, K.; Karthik, K.; Tiwari, R.; Malik, Y. S.; Singh, R. K.; Chaicumpa, W. (2018). Modulation of Dengue/Zika Virus Pathogenicity by Antibody-Dependent Enhancement and Strategies to Protect Against Enhancement in Zika Virus Infection. Frontiers in Immunology 9: 597. PMC 5925603. PMID 29740424. doi:10.3389/fimmu.2018.00597.
- Ruta Kulkarni. Antibody-Dependent Enhancement of Viral Infections // Dynamics of Immune Activation in Viral Diseases. — 2019. — 5 листопада. — С. 9–41. — DOI:.
- Міронов, О. М.; Супотніцький, М. В.; Лебединська, О. В. (2014). Феномен антитілозалежного посилення інфекції у вакцинованих і перехворілих (частина II). Інфекційні хвороби (1). с. 69–80. ISSN 1681-2727. Процитовано 3 липня 2021.
- HAWKES, Royle. www.dpi.nsw.gov.au (англ.). 2020. Процитовано 12 червня 2021.
- Hawkes R.A. Enhancement of the infectivity of arboviruses by specific antisera produced in domestic fowls // Aust. J. Exp. Biol. Med. Sci. 1964. V. 43. P. 465—482.
- Hawkes R.A., Lafferty K.J. The enhancement of virus infectivity by antibody // Virology. 1967. V. 33. P. 250—261.
- Halstead S.B., Chow J., Marchette N.J. Immunologic enhancement of Dengue virus replication // Nat. New Biol. 1973. V. 243. P. 24—26.
- Halstead S.B., Mahalingam P.S., Marovich M.A. et al. Intrinsic antibody-dependent enhancement of microbial infection in macrophages: disease regulation by immune complexes // Lancet Infect. Dis. 2010. V. 10, № 10. P. 712—722.
- Sri Rezeki Hadinegoro, Jose Luis Arredondo-García, Maria Rosario Capeding, Carmen Deseda, Tawee Chotpitayasunondh. Efficacy and Long-Term Safety of a Dengue Vaccine in Regions of Endemic Disease // New England Journal of Medicine. — 2015. — Т. 373, вип. 13 (24 вересня). — С. 1195—1206. — ISSN 0028-4793. — DOI:.
- Saranya Sridhar, Alexander Luedtke, Edith Langevin, Ming Zhu, Matthew Bonaparte. Effect of Dengue Serostatus on Dengue Vaccine Safety and Efficacy // New England Journal of Medicine. — 2018. — Т. 379, вип. 4 (26 липня). — С. 327—340. — ISSN 0028-4793. — DOI:.
- Scott B. Halstead, Philip K. Russell. Protective and immunological behavior of chimeric yellow fever dengue vaccine // Vaccine. — Elsevier, 2016. — Vol. 34, iss. 14 (1 March). — P. 1643—1647. — ISSN 0264-410X. — DOI:.
- Lisa Rosenbaum. Trolleyology and the Dengue Vaccine Dilemma // New England Journal of Medicine. — 2018. — Т. 379, вип. 4 (26 липня). — С. 305—307. — ISSN 1533-4406 0028-4793, 1533-4406. — DOI:.
- WHO | Questions and Answers on Dengue Vaccines
- A year after Dengvaxia: Immunization drops, measles outbreaks soar. Part 1 // Rappler. December 01, 2018
- Post Dengvaxia controversy: Paving the way forward for vaccines, health care. Part 2 // Rappler. December 02, 2018
- Controversy over dengue vaccine causes panic in the Philippines // CGTN. 24 февраля 2018 г.
- Dengue Vaccine | Dengue | CDC (англ.). www.cdc.gov. 23 вересня 2019. Процитовано 12 травня 2020.
- Jon Cohen. New dengue vaccine performs well in large trial, but safety remains key concern // Science. — 2019. — 6 November. — ISSN 1095-9203 0036-8075, 1095-9203. — DOI:.
- Stephen J. Thomas, In-Kyu Yoon. A review of Dengvaxia®: development to deployment // Human Vaccines & Immunotherapeutics. — 2019. — Т. 15, вип. 10 (3 жовтня). — С. 2295–2314. — ISSN 2164-554X 2164-5515, 2164-554X. — DOI:.
- Белло-Джил Д., Манец Р. Использование естественных антиуглеводных антител в терапевтических целях: Обзор // Биохимия. 2015. Т. 80, вып. 7. С. 998—1009.
- Также список вирусов, которые могут вызывать антителозависимое усиление инфекции, приведён на страницах 179—182 в руководстве по проведению доклинических исследований, выпущенном в 2012 году ФГБУ «НЦЭСМП» Минздравсоцразвития России (Меркулов В.А. и др. Руководство по проведению доклинических исследований лекарственных средств (часть 2) / Миронов А. Н. — М. : Гриф и К, 2012. — С. 179—182.
- Феномен антитело-зависимого усиления инфекции у вакцинированных и переболевших. supotnitskiy.ru. Процитовано 14 серпня 2020.
- Coronavirus Resource Center: As coronavirus spreads, many questions and some answers. Updated: May 6, 2020
- WHO Director-General's opening remarks at the media briefing on COVID-19 — 11 March 2020
- ВОЗ объявила о пандемии коронавируса.
- Molecular Mechanism for Antibody-Dependent Enhancement of Coronavirus Entry // Journal of Virology. 11 December 2019.
- Ann M. Arvin, Katja Fink, Michael A. Schmid, Andrea Cathcart, Roberto Spreafico. A perspective on potential antibody-dependent enhancement of SARS-CoV-2 // Nature. — 2020. — 13 July. — P. 1–11. — ISSN 1476-4687. — DOI:.
- Nikolai Eroshenko, Taylor Gill, Marianna K. Keaveney, George M. Church, Jose M. Trevejo. Implications of antibody-dependent enhancement of infection for SARS-CoV-2 countermeasures // Nature Biotechnology. — 2020. — Vol. 38, iss. 7 (1 July). — P. 789–791. — ISSN 1546-1696. — DOI:.
- https://plus.google.com/b/101938688787495382801 (21 квітня 2021). Дивний феномен вакцини Sinovac: країни повідомляють про зростання випадків захворювання після її використання. Велика Епоха (укр.). Процитовано 3 листопада 2021.
- Welle (www.dw.com), Deutsche. Доля привитых среди заразившихся коронавирусом в ФРГ - 5 процентов | DW | 29.10.2021. DW.COM (ru-RU). Процитовано 2 листопада 2021.
- Gavin Koh (22 березня 2020). Faculty Opinions recommendation of SARS CoV subunit vaccine: antibody-mediated neutralisation and enhancement. Faculty Opinions – Post-Publication Peer Review of the Biomedical Literature. Процитовано 30 квітня 2020.
- Hiu-lan, Nancy Leung. Mechanism of antibody-dependent enhancement in severe acute respiratory syndrome coronavirus infection. — The University of Hong Kong Libraries.
- Yip, Ming Leung, Nancy Hiu Cheung, Chung Li, Ping Lee, Horace Hok Daëron, Marc Peiris, Joseph Sriyal Bruzzone, Roberto Jaume, Martial. Antibody-dependent infection of human macrophages by severe acute respiratory syndrome coronavirus.
- Immunodominant SARS Coronavirus Epitopes in Humans Elicited both Enhancing and Neutralizing Effects on Infection in Non-human Primates. dx.doi.org. Процитовано 30 квітня 2020.
- Ming S Yip, Chung Y Cheung, Ping H Li, Roberto Bruzzone, JS Malik Peiris. Investigation of Antibody-Dependent Enhancement (ADE) of SARS coronavirus infection and its role in pathogenesis of SARS // BMC Proceedings. — 2011. — Т. 5, вип. S1 (10 січня). — ISSN 1753-6561. — DOI:.
- Современный взгляд на диагностику, лечение и профилактику инфекционного перитонита кошек / Терехова Ю. О., Цибезов В. В., Верховский О. А., Рахманина Н. А., Елаков А. Л. // VetPharma. 2014. № 2, март.
- Javier A. Jaimes, Gary R. Whittaker. Feline coronavirus: Insights into viral pathogenesis based on the spike protein structure and function // Virology. — 2018. — Vol. 517 (1 April). — P. 108–121. — ISSN 0042-6822. — DOI:.
- Harry Vennema, Amy Poland, Janet Foley, Niels C. Pedersen. Feline Infectious Peritonitis Viruses Arise by Mutation from Endemic Feline Enteric Coronaviruses // Virology. — 1998. — Т. 243, вип. 1 (1 березня). — С. 150—157. — ISSN 0042-6822. — DOI:.
- Beth N. Licitra, Jean K. Millet, Andrew D. Regan, Brian S. Hamilton, Vera D. Rinaldi. Mutation in Spike Protein Cleavage Site and Pathogenesis of Feline Coronavirus - Volume 19, Number 7—July 2013 - Emerging Infectious Diseases journal - CDC. — DOI:.
- H Vennema, R J de Groot, D A Harbour, M Dalderup, T Gruffydd-Jones. Early death after feline infectious peritonitis virus challenge due to recombinant vaccinia virus immunization // Journal of Virology. — 1990. — Т. 64, вип. 3 (15 лютого). — С. 1407—1409. — ISSN 1098-5514 0022-538X, 1098-5514. — DOI:.
- T. Hohdatsu, M. Nakamura, Y. Ishizuka, H. Yamada, H. Koyama. A study on the mechanism of antibody-dependent enhancement of feline infectious peritonitis virus infection in feline macrophages by monoclonal antibodies // Archives of Virology. — 1991. — Т. 120, вип. 3—4 (1 вересня). — С. 207—217. — ISSN 1432-8798 0304-8608, 1432-8798. — DOI:.
- W V Corapi, C W Olsen, F W Scott. Monoclonal antibody analysis of neutralization and antibody-dependent enhancement of feline infectious peritonitis virus. // Journal of Virology. — 1992. — Т. 66, вип. 11 (1 листопада). — С. 6695–6705. — ISSN 0022-538X.
- T. Hohdatsu, M. Yamada, R. Tominaga, K. Makino, K. Kida. Antibody-dependent enhancement of feline infectious peritonitis virus infection in feline alveolar macrophages and human monocyte cell line U937 by serum of cats experimentally or naturally infected with feline coronavirus // The Journal of Veterinary Medical Science. — 1998. — Т. 60, вип. 1 (1 січня). — С. 49–55. — ISSN 0916-7250. — DOI:.
- Tomomi TAKANO, Shinji YAMADA, Tomoyoshi DOKI, Tsutomu HOHDATSU. Pathogenesis of oral type I feline infectious peritonitis virus (FIPV) infection: Antibody-dependent enhancement infection of cats with type I FIPV via the oral route // Journal of Veterinary Medical Science. — 2019. — Т. 81, вип. 6 (15 лютого). — С. 911—915. — ISSN 1347-7439 0916-7250, 1347-7439. — DOI:.
- Francesco Negro. Is antibody-dependent enhancement playing a role in COVID-19 pathogenesis? // Swiss Medical Weekly. — 2020. — 16 квітня. — ISSN 1424-3997. — DOI:.
- Tsutomu Hohdatsu, Hiroshi Yamato, Tasuku Ohkawa, Miyuki Kaneko, Kenji Motokawa. Vaccine efficacy of a cell lysate with recombinant baculovirus-expressed feline infectious peritonitis (FIP) virus nucleocapsid protein against progression of FIP // Veterinary Microbiology. — 2003. — Vol. 97, iss. 1 (2 December). — P. 31–44. — ISSN 0378-1135. — DOI:.
- Ming Shum Yip, Nancy Hiu Lan Leung, Chung Yan Cheung, Ping Hung Li, Horace Hok Yeung Lee. Antibody-dependent infection of human macrophages by severe acute respiratory syndrome coronavirus // Virology Journal. — 2014. — Т. 11, вип. 1 (6 травня). — С. 82. — ISSN 1743-422X. — DOI:.
- Lanjuan Li, Jianer Wo, Junbing Shao, Haihong Zhu, Nanping Wu. SARS-coronavirus replicates in mononuclear cells of peripheral blood (PBMCs) from SARS patients // Journal of Clinical Virology. — 2003. — Т. 28, вип. 3 (1 грудня). — С. 239—244. — ISSN 1386-6532. — DOI:.
- Mamadi Yilla, Brian H. Harcourt, Carole J. Hickman, Marcia McGrew, Azaibi Tamin. SARS-coronavirus replication in human peripheral monocytes/macrophages // Virus Research. — 2005. — Т. 107, вип. 1 (1 січня). — С. 93—101. — ISSN 0168-1702. — DOI:.
- Akiko Iwasaki, Yexin Yang. The potential danger of suboptimal antibody responses in COVID-19 // Nature Reviews Immunology. — Nature Publishing Group, 2020. — 21 April. — ISSN 1474-1741 1474-1733, 1474-1741. — DOI:.
- Immunodominant SARS Coronavirus Epitopes in Humans Elicited both Enhancing and Neutralizing Effects on Infection in Non-human Primates. dx.doi.org. Процитовано 28 вересня 2020.
- Yiu Wing Kam, François Kien, Anjeanette Roberts, Yan Chung Cheung, Elaine W. Lamirande. Antibodies against trimeric S glycoprotein protect hamsters against SARS-CoV challenge despite their capacity to mediate FcgammaRII-dependent entry into B cells in vitro // Vaccine. — 2007. — Т. 25, вип. 4 (8 січня). — С. 729–740. — ISSN 0264-410X. — DOI:.
- Sheng-Fan Wang, Sung-Pin Tseng, Chia-Hung Yen, Jyh-Yuan Yang, Ching-Han Tsao. Antibody-dependent SARS coronavirus infection is mediated by antibodies against spike proteins // Biochemical and Biophysical Research Communications. — 2014. — Т. 451, вип. 2 (22 серпня). — С. 208–214. — ISSN 0006-291X. — DOI:.
- Yushun Wan, Jian Shang, Shihui Sun, Wanbo Tai, Jing Chen. Molecular Mechanism for Antibody-Dependent Enhancement of Coronavirus Entry // Journal of Virology. — . — Т. 94, вип. 5. — ISSN 1098-5514. — DOI:.
- Li Liu, Qiang Wei, Qingqing Lin, Jun Fang, Haibo Wang. Anti–spike IgG causes severe acute lung injury by skewing macrophage responses during acute SARS-CoV infection // JCI Insight. — 2019. — Т. 4, вип. 4 (21 лютого). — ISSN 2379-3708. — DOI:.
- Hana Weingartl, Markus Czub, Stefanie Czub, James Neufeld, Peter Marszal. Immunization with modified vaccinia virus Ankara-based recombinant vaccine against severe acute respiratory syndrome is associated with enhanced hepatitis in ferrets // Journal of Virology. — 2004. — Т. 78, вип. 22 (1 листопада). — С. 12672–12676. — ISSN 0022-538X. — DOI:.
- M. Bolles, D. Deming, K. Long, S. Agnihothram, A. Whitmore. A Double-Inactivated Severe Acute Respiratory Syndrome Coronavirus Vaccine Provides Incomplete Protection in Mice and Induces Increased Eosinophilic Proinflammatory Pulmonary Response upon Challenge // Journal of Virology. — 2011. — Т. 85, вип. 23 (21 вересня). — С. 12201—12215. — ISSN 0022-538X. — DOI:.
- Chien-Te Tseng, Elena Sbrana, Naoko Iwata-Yoshikawa, Patrick C. Newman, Tania Garron. Immunization with SARS Coronavirus Vaccines Leads to Pulmonary Immunopathology on Challenge with the SARS Virus // PLOS One. — Public Library of Science, 2012. — Vol. 7, iss. 4 (20 April). — P. e35421. — ISSN 1932-6203. — DOI:.
- Damon Deming, Timothy Sheahan, Mark Heise, Boyd Yount, Nancy Davis. Vaccine Efficacy in Senescent Mice Challenged with Recombinant SARS-CoV Bearing Epidemic and Zoonotic Spike Variants // PLoS Medicine. — 2006. — Т. 3, вип. 12 (1 грудня). — ISSN 1549-1277. — DOI:.
- Fumihiko Yasui, Chieko Kai, Masahiro Kitabatake, Shingo Inoue, Misako Yoneda. Prior immunization with severe acute respiratory syndrome (SARS)-associated coronavirus (SARS-CoV) nucleocapsid protein causes severe pneumonia in mice infected with SARS-CoV // Journal of Immunology (Baltimore, Md.: 1950). — 2008. — Т. 181, вип. 9 (1 листопада). — С. 6337–6348. — ISSN 1550-6606. — DOI:.
- Katherine V. Houser, Andrew J. Broadbent, Lisa Gretebeck, Leatrice Vogel, Elaine W. Lamirande. Enhanced inflammation in New Zealand white rabbits when MERS-CoV reinfection occurs in the absence of neutralizing antibody // PLoS Pathogens. — 2017. — Т. 13, вип. 8 (17 серпня). — ISSN 1553-7366. — DOI:.
- Anurodh Shankar Agrawal, Xinrong Tao, Abdullah Algaissi, Tania Garron, Krishna Narayanan. Immunization with inactivated Middle East Respiratory Syndrome coronavirus vaccine leads to lung immunopathology on challenge with live virus // Human Vaccines & Immunotherapeutics. — 2016. — Т. 12, вип. 9 (7 червня). — С. 2351—2356. — ISSN 2164-554X 2164-5515, 2164-554X. — DOI:.
- Qidi Wang, Lianfeng Zhang, Kazuhiko Kuwahara, Li Li, Zijie Liu. Immunodominant SARS Coronavirus Epitopes in Humans Elicited both Enhancing and Neutralizing Effects on Infection in Non-human Primates // ACS infectious diseases. — . — Т. 2, вип. 5. — С. 361–376. — ISSN 2373-8227. — DOI:.
- Jaume, Martial Yip, Ming S. Cheung, Chung Y. Leung, Hiu L. Li, Ping H. Kien, Francois Dutry, Isabelle Callendret, Benoît Escriou, Nicolas Altmeyer, Ralf Nal, Beatrice Daëron, Marc Bruzzone, Roberto Peiris, J. S. Malik. Anti-Severe Acute Respiratory Syndrome Coronavirus Spike Antibodies Trigger Infection of Human Immune Cells via a pH- and Cysteine Protease-Independent FcγR Pathway ▿. — American Society for Microbiology.
- F. F. Yuan, J. Tanner, P. K. S. Chan, S. Biffin, W. B. Dyer. Influence of FcgammaRIIA and MBL polymorphisms on severe acute respiratory syndrome // Tissue Antigens. — 2005. — Т. 66, вип. 4 (1 жовтня). — С. 291—296. — ISSN 1399-0039 0001-2815, 1399-0039. — DOI:.
- Stylianos Bournazos, Aaron Gupta, Jeffrey V. Ravetch. The role of IgG Fc receptors in antibody-dependent enhancement // Nature Reviews. Immunology. — 2020. — 11 серпня. — ISSN 1474-1741. — DOI:.
- W V Corapi, C W Olsen, F W Scott. Monoclonal antibody analysis of neutralization and antibody-dependent enhancement of feline infectious peritonitis virus. // Journal of Virology. — 1992. — Т. 66, вип. 11 (15 лютого). — С. 6695–6705. — ISSN 1098-5514 0022-538X, 1098-5514. — DOI:.
- Darrell Ricke, Robert W. Malone. Medical Countermeasures Analysis of 2019-nCoV and Vaccine Risks for Antibody-Dependent Enhancement (ADE) // SSRN Electronic Journal. — 2020. — 15 лютого. — ISSN 1556-5068. — DOI:.
- Wladek Minor, Ivan Shabalin (20 лютого 2020). Faculty Opinions recommendation of Cryo-EM structure of the 2019-nCoV spike in the prefusion conformation. Faculty Opinions – Post-Publication Peer Review of the Biomedical Literature. Процитовано 30 квітня 2020.
- Proteolytic Cleavage of the SARS-CoV-2 Spike Protein and the Role of the Novel S1/S2 Site // iScience. — 2020. — Vol. 23, iss. 6 (26 June). — P. 101212. — ISSN 2589-0042. — DOI:.
- Ting Gao, Mingdong Hu, Xiaopeng Zhang, Hongzhen Li, Lin Zhu. Highly pathogenic coronavirus N protein aggravates lung injury by MASP-2-mediated complement over-activation // medRxiv. — 2020. — 18 June. — P. 2020.03.29.20041962. — DOI:.
- Jeremia M. Coish, Adam J. MacNeil. Out of the frying pan and into the fire? Due diligence warranted for ADE in COVID-19 // Microbes and Infection. — 2020. — 24 червня. — ISSN 1286-4579. — DOI:.
- Yajing Fu, Yuanxiong Cheng, Yuntao Wu. Understanding SARS-CoV-2-Mediated Inflammatory Responses: From Mechanisms to Potential Therapeutic Tools // Virologica Sinica. — 2020. — Vol. 35, iss. 3 (1 June). — P. 266–271. — ISSN 1995-820X. — DOI:.
- Kumaragurubaran Karthik, Tuticorin Maragatham Alagesan Senthilkumar, Shanmugasundaram Udhayavel, Gopal Dhinakar Raj. Role of antibody-dependent enhancement (ADE) in the virulence of SARS-CoV-2 and its mitigation strategies for the development of vaccines and immunotherapies to counter COVID-19 // Human Vaccines & Immunotherapeutics. — 2020. — 26 серпня. — С. 1–6. — ISSN 2164-554X. — DOI:.
- Maryse Cloutier, Madhuparna Nandi, Awais Ullah Ihsan, Hugues Allard Chamard, Subburaj Ilangumaran. ADE and hyperinflammation in SARS-CoV2 infection- comparison with dengue hemorrhagic fever and feline infectious peritonitis // Cytokine. — 2020. — Т. 136 (1 грудня). — С. 155256. — ISSN 1043-4666. — DOI:.
- Институт молекулярной биологии им. В.А. Энгельгардта РАН, Ю.Д. Нечипуренко, А.А. Анашкина, Институт молекулярной биологии им. В.А. Энгельгардта РАН, О.В. Матвеева. ИЗМЕНЕНИЕ АНТИГЕННЫХ ДЕТЕРМИНАНТ S-БЕЛКА ВИРУСА SARS-COV-2 КАК ВОЗМОЖНАЯ ПРИЧИНА АНТИТЕЛОЗАВИСИМОГО УСИЛЕНИЯ ИНФЕКЦИИ И ЦИТОКИНОВОГО ШТОРМА // Биофизика. — 2020. — Т. 65, вип. 4 (15 лютого). — С. 824–832. — DOI:.
- Wen Shi Lee, Adam K. Wheatley, Stephen J. Kent, Brandon J. DeKosky. Antibody-dependent enhancement and SARS-CoV-2 vaccines and therapies // Nature Microbiology. — 2020. — Vol. 5, iss. 10 (1 October). — P. 1185–1191. — ISSN 2058-5276. — DOI:.
- Yu A. Desheva, A. S. Mamontov, P. G. Nazarov. Contribution of antibody-dependent enhancement to the pathogenesis of coronavirus infections // AIMS Allergy and Immunology. — 2020. — Vol. 4, iss. 3 (15 February). — P. 50. — DOI:.
- Ватутин Н. Т., Ещенко Е. В. Лимфопения: основные причины развития // Архивъ внутренней медицины. — 2016. — № 2(28). — С. 22-27.
- Ajai A. Dandekar, Stanley Perlman. Immunopathogenesis of coronavirus infections: implications for SARS // Nature Reviews Immunology. — 2005. — Т. 5, вип. 12 (1 грудня). — С. 917–927. — ISSN 1474-1741 1474-1733, 1474-1741. — DOI:.
- Linqi Zhang, Fengwen Zhang, Wenjie Yu, Tian He, Jian Yu. Antibody responses against SARS coronavirus are correlated with disease outcome of infected individuals // Journal of Medical Virology. — 2005. — Т. 78, вип. 1 (15 лютого). — С. 1—8. — ISSN 1096-9071 0146-6615, 1096-9071. — DOI:.
- Nelson Lee, P. K. S. Chan, Margaret Ip, Eric Wong, Jenny Ho. Anti-SARS-CoV IgG response in relation to disease severity of severe acute respiratory syndrome // Journal of Clinical Virology: The Official Publication of the Pan American Society for Clinical Virology. — 2006. — Т. 35, вип. 2 (1 лютого). — С. 179–184. — ISSN 1386-6532. — DOI:.
- Mei-Shang Ho, Wei-Ju Chen, Hour-Young Chen, Szu-Fong Lin, Min-Chin Wang. Neutralizing Antibody Response and SARS Severity // Emerging Infectious Diseases. — 2005. — Т. 11, вип. 11 (1 листопада). — С. 1730–1737. — ISSN 1080-6040. — DOI:.
- Jiang Gu, Clive R. Taylor. Acute Immunodeficiency, Multiple Organ Injury, and the Pathogenesis of SARS // Applied Immunohistochemistry & Molecular Morphology. — 2003. — 1 грудня. — С. 281—282. — ISSN 1541-2016. — DOI:.
- Hangping Yao, Xiangyun Lu, Qiong Chen, Kaijin Xu, Yu Chen, Linfang Cheng (19 квітня 2020). Patient-derived mutations impact pathogenicity of SARS-CoV-2. dx.doi.org. Процитовано 30 квітня 2020.
- Yasunori Watanabe, Joel D. Allen, Daniel Wrapp, Jason S. McLellan, Max Crispin (28 березня 2020). Site-specific analysis of the SARS-CoV-2 glycan shield. dx.doi.org. Процитовано 11 серпня 2020.
- Peter L. Nara, Robert R. Garrity, Jaap Goudsmit. Neutralization of HIV‐1: a paradox of humoral proportions // The FASEB Journal. — 1991. — Т. 5, вип. 10 (1 липня). — С. 2437–2455. — ISSN 1530-6860 0892-6638, 1530-6860. — DOI:.
- O. Narayan, D. E. Griffin, J. E. Clements. Virus Mutation during 'Slow Infection': Temporal Development and Characterization of Mutants of Visna Virus recovered from Sheep // Journal of General Virology. — 1978. — Т. 41, вип. 2 (1 листопада). — С. 343–352. — ISSN 1465-2099 0022-1317, 1465-2099. — DOI:.
- Y. Kono, K. Kobayashi, Y. Fukunaga. Serological comparison among various strains of equine infectious anemia virus // Archiv f�r die gesamte Virusforschung. — 1971. — Т. 34, вип. 3 (1 вересня). — С. 202–208. — ISSN 1432-8798 0304-8608, 1432-8798. — DOI:.
- Christopher P. Locher, Robert M. Grant, Eric A. Collisson, Gustavo Reyes-Teran, Tarek Elbeik. Short Communication Antibody and Cellular Immune Responses in Breakthrough Infection Subjects after HIV Type 1 Glycoprotein 120 Vaccination // AIDS Research and Human Retroviruses. — 1999. — Т. 15, вип. 18 (10 грудня). — С. 1685–1689. — ISSN 1931-8405 0889-2229, 1931-8405. — DOI:.
- Natasha Larke, Eung-Jun Im, Ralf Wagner, Carolyn Williamson, Anna-Lise Williamson. Combined single-clade candidate HIV-1 vaccines induce T cell responses limited by multiple forms ofin vivo immune interference // European Journal of Immunology. — 2007. — Т. 37, вип. 2 (1 лютого). — С. 566–577. — ISSN 1521-4141 0014-2980, 1521-4141. — DOI:.
- S. MULLER, H. WANG, G. J. SILVERMAN, G. BRAMLET, N. HAIGWOOD. B-Cell Abnormalities in AIDS: Stable and Clonally-Restricted Antibody Response in HIV-1 Infection // Scandinavian Journal of Immunology. — 1993. — Т. 38, вип. 4 (1 жовтня). — С. 327–334. — ISSN 1365-3083 0300-9475, 1365-3083. — DOI:.
- Jop Vrieze. First malaria vaccine rolled out in Africa—despite limited efficacy and nagging safety concerns // Science. — 2019. — 26 листопада. — ISSN 1095-9203 0036-8075, 1095-9203. — DOI:.
- Patrick E. Duffy, J. Patrick Gorres. Malaria vaccines since 2000: progress, priorities, products // npj Vaccines. — 2020. — Vol. 5, iss. 1 (9 June). — P. 1–9. — ISSN 2059-0105. — DOI:.
- M. F. Good, Y. Zevering, J. Currier, J. Bilsborough. 'Original antigenic sin', T cell memory, and malaria sporozoite immunity: an hypothesis for immune evasion // Parasite Immunology. — 1993. — Т. 15, вип. 4 (1 квітня). — С. 187–193. — ISSN 0141-9838. — DOI:.
- E. M. Riley. The role of MHC- and non-MHC-associated genes in determining the human immune response to malaria antigens // Parasitology. — 1996. — Т. 112 Suppl (15 лютого). — С. S39–51. — ISSN 0031-1820.
- Richard J. Pleass, Solabomi A. Ogun, David H. McGuinness, Jan G. J. van de Winkel, Anthony A. Holder. Novel antimalarial antibodies highlight the importance of the antibody Fc region in mediating protection // Blood. — 2003. — Vol. 102, iss. 13 (15 December). — P. 4424–4430. — ISSN 0006-4971. — DOI:.
- Jiraprapa Wipasa, Huji Xu, Xueqin Liu, Chakrit Hirunpetcharat, Anthony Stowers. Effect of Plasmodium yoelii Exposure on Vaccination with the 19-Kilodalton Carboxyl Terminus of Merozoite Surface Protein 1 and Vice Versa and Implications for the Application of a Human Malaria Vaccine // Infection and Immunity. — 2009. — Vol. 77, iss. 2 (1 February). — P. 817–824. — ISSN 1098-5522 0019-9567, 1098-5522. — DOI:.
- Complement and Antibody-mediated Enhancement of Red Blood Cell Invasion and Growth of Malaria Parasites // EBioMedicine. — 2016. — Vol. 9 (1 July). — P. 207–216. — ISSN 2352-3964. — DOI:.
- Laurent Rénia, Yun Shan Goh. Malaria Parasites: The Great Escape // Frontiers in Immunology. — 2016. — Т. 7 (7 листопада). — ISSN 1664-3224. — DOI:.
Література
- Феномен антителозависимого усиления инфекции при доклиническом изучении иммунобиологических лекарственных препаратов // Руководство по проведению доклинических исследований лекарственных средств (иммунобиологические лекарственные препараты): В 2 частях. Ч. 2 / Под ред. А. Н. Миронова; ФГБУ «НЦЭМСП» Минздравсоцразвития России. — М. : Гриф и К, 2012. — С. 177—185.
- Богадельников І. В., Крюгер Е. А. Останнє слово буде за мікробами? // Клінічна інфектологія та паразитологія. — 2012. — № 2. — С. 4-12.
- Богадельников И. В. Микроорганизмы — властители эволюции / Мед. акад. им. С. И. Георгиевского, ФГАУВО «КФУ им. В. И. Вернадского». — Симферополь : Н. Орiанда, 2016. — 150 с. — 300 прим. — ISBN 978-5-9907469-8-5.
Посилання
- Ржешевский А. В. Віруси і людина. Протистояння довжиною в тисячоліття // Biomolecula.ru. 8 листопада 2015.
- Анна Урманцева, Марія Недюк. Така корона: вакцина від COVID-19 може погіршити захворювання: Фахівці побоюються розвитку так званого синдрому антитіло-залежного посилення інфекції // Вісті: Наука. 3 березня 2020.
- У коронавірусу знайшли нову небезпечну мутацію // Lenta.ru. 6 травня 2020. (Антитілозалежне посилення інфекції як основа гіпотез про механізми мутацій) — Spike mutation pipeline reveals the emergence of a more transmissible form of SARS-CoV-2
- Topic 5 — Human Immune Pathways: Theory About Antibody Dependent Enhancement (Presented by: Dr. Nor Omaima Harun) // UMTMOOC. 21 грудня 2017 р. (відео)(англ.)
- Antibody Dependent Enhancement in Dengue Infection / / Zayan Mahmooth 21 серпня 2017 р. (відео)(англ.)
- CV Central — 5/4/2020 «Transmissibility, Mutations, and Immune Enhancement» // Tom Kawczynski. 4 травня 2020 р.(англ.)
- Ольга Матвєєва: «я б не лила плазму всім підряд» // Голос Америки. 6 травня 2020 р. (Мікробіолог — про те, чи рівні антитіла імунітету, про надійність тестів і про небезпеку антитілозалежного посилення інфекції)
- «Керівництво з проведення доклінічних досліджень лікарських засобів (імунобіологічні лікарські препарати)» у двох частинах. У розділі "Феномен антитілозалежного посилення інфекції при доклінічному вивченні імунобіологічних лікарських препаратів"наведено докладні інструкції тестування вакцин.
