Дасплетозавр
Дасплетозавр (Daspletosaurus torosus Russell, 1970 typus)[1] (лат. Daspletosaurus, від дав.-гр. δασπλῆτις «грізна», епітет Ериній або Гекати, і σαῦρος «ящірка») — рід м'ясоїдних динозаврів крейдового періоду родини тиранозаврид, який жив на заході Північної Америки. Рід Дасплетозаври мав два види. Скам'янілості раннього виду, D. torosus, були знайдені в провінції Альберта, тоді як скам'янілості пізнішого другого виду, D. horneri, були знайдені лише в Монтані. Можливий третій вид, також з Альберти, досі чекає на офіційну ідентифікацію.
| Дасплетозавр | |
|---|---|
 | |
| Реконструкція кістяка D. torosus. | |
| Біологічна класифікація | |
| Царство: | Тварини (Animalia) |
| Тип: | Хордові (Chordata) |
| Клада: | Динозаври (Dinosauria) |
| Ряд: | Ящеротазові (Saurischia) |
| Підряд: | Тероподи (Theropoda) |
| Родина: | †Тиранозаврові (Tyrannosauridae) |
| Підродина: | †Tyrannosaurinae |
| Рід: | †Дасплетозавр (Daspletosaurus) Russell, 1970 |
| Типовий вид | |
| †Daspletosaurus torosus Russell, 1970 | |
| Види | |
Дасплетозавр близько споріднений зі значно більшим і більш пізнім тиранозавридом Тиранозавром Рексом. Як і більшість тиранозаврид, Дасплетозавр був багатотонним двоногим хижаком, озброєним безліччю великих гострих зубів. Дасплетозавр мав маленькі передні кінцівки, типові для тираннозаврид, хоча вони були пропорційно довші, ніж в інших видів.
Як найсильніший хижак, Дасплетозавр був на вершині харчового ланцюга, напевне полюючи на великих динозаврів, таких як Цератопсид, Центрозавр, Гадрозавр, Гіпакрозавр. На деяких територіях Дасплетозавр співіснував з іншим тиранозавридом, Горгозавром, хоча є деякі докази відмінностей в способі життя між цими двома видами. Хоча скам'янілості Дасплетозаврів рідкісні, порівнюючи з іншими тиранозавридами, доступні зразки дозволяють провести деякий аналіз з біології цих тварин, включно з соціальною поведінкою, способом харчування і життєвим циклом.
Опис
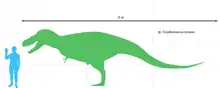
Будучи дуже великим відносно сучасних хижаків, Дасплетозавр не був найбільшим з тираннозаврид. Дорослі особини могли досягати довжини 8-9 метрів від кінчика носа до кінчика хвоста[2]. Приблизна оцінка ваги динозавра — близько 2,5 тонн[2][3][4], варіює від 1,8[5] до 3,8 тонни[6].

Череп Дасплетозавра був масивним, міг досягати понад 1 метр у довжину[2]. Кістки черепа були міцними, носові кістки були щільно зрощені для збільшення сили укусу. Великі отвори в черепі знижували його вагу. Дорослі Дасплетозаври мали близько 60-ти зубів, вони були не дуже довгими, зате широкими, на відміну від зубів Карнозаврів, які були тонкими як леза. На передщелепній кістці, зуби розташовувалися в D-подібному перетині, як і в інших тиранозаврів. Унікальним череп Дасплетозавра робила дуже груба поверхня щелепної кістки та потужні гребені, які проходили навколо очниць, слізних кісток, посторбітальних і виличних. Очниці мали овальну форму, в той час як в інших тираннозаврид очниці нагадували викривлену букву В[7][8][9].
Дасплетозавр мав ту ж саму форму тіла, що і всі тираннозавриди, з короткою S-подібною шиєю, що підтримує масивний череп. Пересувався він на двох ногах, що закінчувалися чотирма пальцями, але перший палець був рудиментарним і не стикався з землею. Передні лапи, навпаки, були дуже короткими, хоча щодо розмірів тіла він має найдовші лапи серед тиранозаврів. Довгий хвіст служив балансиром, центр ваги припадав на тазову область[2][9].
Вивчення черепів D. horneri дозволяє припустити, що на відміну від багатьох зображень тиранозаврид, у цього роду і напевне в інших тиранозаврид були відсутні губи. Порівняння черепа цього виду з сучасними крокодилами показує таку саму складчасту будову кісток, яка свідчить про наявність великих пласких лускоподібних пластин, протиставлених губам. Через те, що шорсткі тканини спускаються аж до лінії зубів, було припущено, що зуби Дасплетозавра не були вкриті губами, тому що там не було місця для таких м'яких тканин.
 Дасплетозавр
Дасплетозавр
Відкриття виду і створення назви
Типовий зразок Daspletosaurus torosus (Канадський музей природи 8506) — це неповний скелет, який охоплює череп, плече, передню кінцівку, таз, стегнову кістку і всі хребці з шиї та тулуба, а також перші одинадцять хребців хвоста. Його було знайдено в 1921 році, поблизу Стіввілля, провінція Альберта, Чарльзом Мортрамом Стернбергом, який думав, що він знайшов новий вид Горгозавра. Так вважали аж до 1970 року, коли зразок був повністю описаний Дейлом Расселом, який дав йому родову назву Дасплетозавр («жахаюча ящірка»). Основна назва Daspletosaurus torosus, слово torosus в латинській мові означає «мускулястий» або «дужий». Крім основного зразка, є лише один добре відомий зразок (Королівський Тиррелівський музей палеонтології 2001.36.1) — відносно повний скелет, знайдений в 2001 році. Обидва зразки були знайдені на території Формації Старого у Річковій Групі Джудіт провінції Альберта. Формація Старого утворилася під час пізнього крейдового періоду від 77 до 76 мільйонів років тому. Зразок з молодшої Формації Каньйону Підкови було перейменовано на Albertosaurus sarcophagus.
Пов'язані види
Два чи три додаткові види були внесені до роду Дасплетозаври, хоча аж до 2007 року ні один з цих видів не отримав належного опису чи наукового імені. Тим часом, всі вони позначені як Дасплетозаври, хоча це не означає, що вони є одним видом.
Разом з голотипом, Расселл позначив зразок, знайдений Барнумом Брауном в 1913 році як паратип виду D. torosus. Цей зразок (Американський музей природознавства 5438) складається з частин задньої ноги, тазу і деяких тазових хребців. Його було знайдено в Формації Парку Динозаврів в Альберті. Формація Парку Динозаврів спочатку була відома як Верхня Формація Старого і належить до проміжку часу між 76,5 і 74,8 мільйонів років тому. Скам'янілі останки Дасплетозаврів відомі з середньої частини верхньої секції формації, між 75.6 і 75.0 мільйонів років тому. В 1914 році Браун знайшов майже повний скелет і череп; 40 років потому Американський музей природознавства продав цей зразок в Музей природної історії ім. Філда в Чикаго. Його було вибрано для виставки в Чикаго і багато років його визначали як Albertosaurus libratus, але після того як кілька властивостей черепа були виліплені з штучних матеріалів, включно з більшістю зубів, зразок (Польовий музей природничої історії PR308) було перейменовано на Daspletosaurus torosus Томасом Карром в 1999 році. Як підсумок, вісім зразків було знайдено відтоді в Формації Парку Динозаврів, більшість з них в межах Провінційного Парку Динозаврів. Філ Каррі вважає, що зразки з Парку Динозаврів являють собою новий вид Дасплетозавра, який відрізняється деякими особливостями черепа. Зображення цього нового виду були опубліковані, але він досі чекає на опублікування імені та повного опису.
Новий зразок тиранозаврид (Музей природничої історії Оклахоми 10131), включно з фрагментами черепа, ребрами і частинами задньої кінцівки, було знайдено в Нью-Мексико в 1990 році і визначено як тепер неіснуючий рід Aublysodon. Багато пізніших авторів заново визначали цей зразок, разом з кількома іншими з Нью-Мексико, як ще один неназваний вид Дасплетозавра. Проте дослідження, опубліковані в 2010 році, показують, що цей вид, з Каньйону Мисливця в Формації Кіртланда, насправді є більш примітивним тиранозавридом і його було класифіковано як рід Bistahieversor. Вчені досі сперечаються про можливий вік Формації Кіртланда.
В 1992 році Джек Хорнер і його колеги опублікували передчасну доповідь про тиранозаврида з верхніх частин Другої Медичної Формації в Монтані, якого було інтерпретовано як перехідний вид між Дасплетозавром і пізнішим Тиранозавром. Каррі (2003) заявляв, що Тиранозаврид з Другої Медичної Формації, згаданий Хорнером (1992), може бути неназваним третім видом Дасплетозавра. Інший неповний скелет було знайдено в верхній частині Другої Медичної Формації в 2001 році, разом з останками молодого Гадрозавра в своїй черевній порожнині. Цей зразок було визначено як Дасплетозавр, але точний вид вказано не було. Останки як мінімум трьох Дасплетозаврів були описані Каррі в покладах кісток на території Другої Медичної Формації (2005), автори заявляли, що цей скам'янілий матеріал напевне є тим самим неназваним видом, який згадував Хорнер (1992), але попереджали, що необхідно продовжувати дослідження і опис, перш ніж цей вид буде визначено з впевненістю. В 2017 році, таксон з Другої Медичної Формації було названо як новий вид D. horneri.
Класифікація і систематика
Дасплетозавр належить до підродини тиранозаврин родини тиранозаврид, разом з Тарбозавром, Тиранозавром і Аліорамом. Тварини з цієї підродини більш близькі до Тиранозавра, ніж до Альбертозавра і відомі, за виключенням Аліорама, через їхню міцну будову тіла з пропорційно більшими черепами і довшими стегнами ніж в іншій підродині Альбертозаврин.
Дасплетозавра звичайно розглядають як близького родича Тиранозавр Рекса, або навіть його прямого предка в результаті анагенезу. Грегорі Пол призначив D. torosus в рід Тиранозаври, створивши нову комбінацію Tyrannosaurus torosus, але це не було прийнято іншими вченими. Багато дослідників вірили, що Тарбозавр і Тиранозавр є близькими родичами або навіть одним родом, з Дасплетозавром як більш базовим родичем. З іншого боку, Філ Каррі і його колеги вважали Дасплетозавра більш близьким до Тарбозавра і інших азійських тиранозаврид, таких як Аліорам, ніж до північноамериканського Тиранозавра. Систематика (еволюційні зв'язки) Дасплетозавра може стати яснішою, коли всі види будуть описані.
Палеобіологія
Чуття
Є докази того, що D. horneri мав покривні органи чуття, які напевне використовувалися для спілкування, модуляції точних рухів щелеп, відчуття температури та виявлення здобичі. Великі пласкі лускоподібні пластини могли додатково захищати морду тварини під час полювання на здобич і внутрішньовидових поєдинків.
Співіснування з Горгозавром
В Північній Америці Дасплетозавр був сучасником тиранозаврида з групи альбертозаврин Горгозавра. Це один з небагатьох прикладів співіснування двох родів тиранозаврид. У сучасних хижаків, хижаки одного розміру розділені в різні екологічні ніші анатомічними, поведінковими чи географічними відмінностями, які обмежують суперництво. Декілька досліджень пробували пояснити різницю в способі життя між Дасплетозавром і Горгозавром.
Дейл Расселл висував гіпотезу, що більш поширений Горгозавр з легкою будовою тіла міг полювати на більш поширених гадрозаврів, в той час як більш міцний і менш поширений Дасплетозавр міг спеціалізуватися на менш поширених, але краще захищених цератопсидах, яких було важче вполювати. Проте, зразок Дасплетозавра (Музей Старого Шляху 200) з Другої Медичної Формації має перетравлені останки молодого гадрозавра в області живота. Більші і ширші морди тиранозаврин, таких як Дасплетозавр, були механічно сильнішими, ніж нижчі морди альбертозаврин, таких як Горгозавр, хоча сила зубів в цих двох груп схожа. Це може означати різницю в механіці харчування чи в дієті.
Інші автори висували припущення, що суперництво було обмежене географічним поділом. На відміну від деяких інших груп динозаврів, схоже тут не було взаємозв'язку з віддаленістю від моря. Ні Дасплетозавр, ні Горгозавр не був більш поширеним на вищих чи нижчих підвищеннях ніж інший. Проте, в той час як тут є деяке перекривання, схоже що Горгозаври були більш поширені в північних широтах, в той час як види Дасплетозаврів були більш поширені на півдні. Таку саму схему видно і в інших груп динозаврів. Хасмозаврини цератопсиди й гадрозаври також є більш поширеними в Другій Медичній Формації та в південнозахідній Північній Америці. Томас Холтз висував припущення, що ця схема свідчить про спільні екологічні особливості між тиранозавринами, хасмозавринами і гадрозавринами. Холтз відзначав що, тиранозаврини, такі як Тиранозавр Рекс, гадрозаврини і хасмозаврини, такі як Трицератопс, були широко поширені в західній Північній Америці, тоді як альбертозаврини і центрозаврини вимерли, і ламбеозаврини були дуже рідкісними.
Соціальна поведінка

Новий зразок Дасплетозавра з Парку Динозаврів (Королівський Тиррелівський музей палеонтології 94.143.1) має сліди від укусів на морді, які були заподіяні іншим тиранозавром. Сліди від укусів пізніше загоїлися, що свідчить про те, що тварина пережила укус. Дорослий Дасплетозавр з Парку Динозаврів (Королівський Тиррелівський музей палеонтології 85.62.1) також має сліди від укусів тиранозавра, що доводить, що атаки в лице не обмежувалися молодими тваринами. Хоча можливо, що укуси могли бути приписані іншим видам, внутрішньовидова агресія, включно з укусами в лице, дуже поширена серед хижаків. Укуси в лице бачили в інших тиранозаврів, таких як Горгозавр і Тиранозавр, також в інших родів тероподів, таких як Сінраптор і Заврорнітолестес. Даррен Тенк і Філ Каррі висували припущення, що укуси були заподіяні через внутрішньовидове суперництво за територію чи ресурси, або перевагу в соціальній групі.
Ознаки того, що Дасплетозаври жили соціальними групами, знайдені в покладах кісток Другої Медичної Формації в Монтані. Поклади кісток містять в собі останки трьох Дасплетозаврів, включно з великою дорослою особиною, малим підлітком і ще однією особиною проміжного розміру. Як мінімум п'ять гадрозаврів збереглися на тому самому місці. Геологічні ознаки свідчать про те, що останки не були принесені разом течією річки, але означають, що всі тварини були похоронені одночасно на одному місці. Останки гадрозаврів розкидані і мають багато слідів від зубів тиранозаврів, що свідчить про те, що Дасплетозаври годувалися гадрозаврами в момент смерті. Причина їх смерті невідома. Каррі робив припущення, що Дасплетозаври складали зграю, хоча це не можна визначити з впевненістю. Інші вчені скептично відносяться до ознак того, що в Дасплетозаврів і інших великих тероподів були соціальні групи.
Брайан Роач і Деніел Брінкман висували припущення, що соціальна взаємодія Дасплетозаврів була більш схожа на ту, яка є в сучасного дракона Комодо, де не соціальні особини оточують тушу тварини, часто атакуючи і навіть поїдаючи одне одного. Докази каннібалізму в Дасплетозаврів були опубліковані в 2015 році.
Життєвий циклу
Палеонтолог Грегорі Еріксон і його колеги досліджували розвиток і життєвий цикл тиранозаврид. Аналіз гістології кісток може встановити вік зразка на момент смерті. Швидкість росту можна вивчати, якщо вік різних особин порівняти з їхнім розміром за допомогою графіку. Еріксон довів, що після довгого життя як підлітки, тиранозаври мали величезні стрибки росту протягом приблизно чотирьох років посередині їхнього життя. Коли фаза швидкого росту закінчувалася статевою зрілістю, у дорослих тварин ріст сильно уповільнювався. Еріксон вивчав Дасплетозавра тільки з Формації Парку Динозаврів, але ці зразки показали таку саму схему. Порівняно з альбертозавринами, Дасплетозаври показували більшу швидкість росту під час періоду швидкого розвитку через більшу вагу дорослих особин. Максимальна швидкість росту в Дасплетозаврів була 180 кілограмів (400 фунтів) на рік, виходячи з оцінки ваги 1800 кілограмів (2 тонни) у дорослих. Інші автори висували припущення про більшу вагу дорослих у Дасплетозаврів; це б змінило величину швидкості росту, але не загальну схему.
Створивши таблицю з кількістю зразків на кожну вікову групу, Еріксон і його колеги змогли отримати висновки про життєвий цикл популяції Альбертозаврів. Їхній аналіз показав, що молодняк знаходили рідко серед скам'янілостей, в той час як особини в фазі швидкого росту і дорослі траплялися набагато частіше. Можливо це було упереджене ставлення, але Еріксон висував припущення, що різниця з'явилася через низьку смертність серед підлітків однакового розміру, що можна побачити у деяких сучасних великих ссавців, таких як слони. Ця низька смертність могла виникнути внаслідок нестачі хижаків, бо тиранозаври випереджали всіх хижаків тієї епохи в розмірі у віці двох років. Палеонтологи не знайшли достатньо останків Дасплетозаврів для схожого аналізу, проте Еріксон відзначав, що та сама загальна тенденція напевне працює і тут.
Дослідження, проведені у 2009 році, знайшли ознаки того, що на щелепах різних зразків Дасплетозаврів була інфекція Trichomonas gallinae.
Палеоекологія
Всі відомі скам'янілі останки Дасплетозавра були знайдені в формаціях, які відносяться до пізнього крейдового періоду, від 77 до 74 мільйонів років тому. Починаючи з середини крейдового періоду, Північна Америка була поділена на дві частини Західним Внутрішнім Морем, з більшою частиною Монтани йАльберти під водою. Проте, підйом Скелястих Гір в гороутворенні Ларамід на заході, який почався в часи Дасплетозаврів, змусив море відступити на схід і південь. Річки спускалися з гір і впадали в море, несучи з собою осадові породи, які створили Другу Медичну Формацію, Річкову Групу Джудіт та інші осадові формації в цьому регіоні. Близько 73 мільйонів років тому, море знову почало рухатися на захід і північ, і весь регіон був вкритий Морем Ведмежої Лапи.
Дасплетозаври жили на безмежних затоплених рівнинах, уздовж західного берега внутрішнього моря. Великі річки наводнювали землю, час від часу затоплюючи й укриваючи регіон новими осадовими породами. Коли води було багато, регіон міг підтримувати багато рослинного і тваринного життя, але періодичні засухи також уражали регіон, які спричиняли велику смертність, що можна побачити в покладах кісток, знайдених в осадових породах з Другої Медичної Формації та річки Джудіт, включно з покладами кісток Дасплетозавра. Схожі умови існують сьогодні в Східній Африці. Вулканічні виверження на заході періодично вкривали регіон попелом, також спричиняючи велику смертність, і одночасно удобрюючи ґрунт для майбутнього росту рослин. Ці поклади попелу також дозволили провести точне радіоізотопне датування. Рівень моря, який часто змінювався, також утворив багато інших середовищ в різні часи в різних ділянках Річкової Групи Джудіт, включно з далекими й близькими до берега природними середовищами в прибережній смузі, прибережними дощовими землями, дельтами й лагунами, на додачу до внутрішніх затоплених рівнин. Друга Медична Формація була розташована на підвищеннях у внутрішніх регіонах у порівнянні з двома іншими формаціями.
Прекрасні скам'янілості хребетних тварин з Другої Медичної Формації та гірських порід річки Джудіт виникли внаслідок поєднання багатого тваринного життя, періодичних природних лих та відкладення великої кількості осадових порід. Багато прісноводних і дельтових видів риби є присутніми, включно з акулами, скатами, осетрами та іншими. Річкова Група Джудіт містить в собі останки багатьох водних амфібій і рептилій, включно з жабами, саламандрами, черепахами, Хампсозаврами й крокодилами. Наземні ящірки, включно з варанами й алігатороподібними ящірками, також були виявлені. Птерозаври Аждарчиди й птахи, такі як Апаторніс і Авізавр, літали над головою, у той час як кілька видів ссавців співіснували з Дасплетозавром і іншими видами динозаврів в різних формаціях, які утворювали клин річки Джудіт.
В Формації Старого (геологічний еквівалент формації річки Джудіт), Daspletosaurus torosus міг полювати на вид гадрозавра Brachylophosaurus canadensis, цератопсин Coronosaurus brinkmani і Albertaceratops nesmoi, пахіцефалозаврів, орнітомімідів, теризинозаврів і можливо анкілозаврів. Інші хижаки охоплювали троодонтид, овірапторозаврів, дромеозаврида Заврорнітолестеса і можливо тиранозавра з групи альбертозаврин (рід на цей момент невідомий). Молодший Парк Динозаврів і Друга Медична Формація мали фауни, схожі на фауну Формації Старого, особливо Парк Динозаврів, який мав дуже багато різновидів динозаврів. Альбертозаврин Горгозавр жив поруч з неназваними видами Дасплетозавра в природних середовищах Парку Динозаврів і верхніх частин Другої Медичної Формації. Молоді тиранозаври могли займати проміжні положення між дорослими тиранозаврами й меншими тероподами, які відрізнялися від двох груп за вагою. Останки Заврорнітолестеса були знайдені в Формації Парку Динозаврів, які мали сліди зубів, які залишилися від укусу молодого тиранозавра, можливо Дасплетозавра.
Примітки
- Бейлі Дж., Седдон Т. {{{Заголовок}}} / І. Б. Шустова. — М., 1995. — С. 39, 100-101, 116. — ISBN 5-7519-0097-9.
- Russell, Dale A. (1970). Tyrannosaurs from the Late Cretaceous of western Canada. National Museum of Natural Sciences Publications in Paleontology 1: 1–34.
- Paul, Gregory S. (1 988). Predatory Dinosaurs of the World. New York: Simon & Schuster. с. 464pp. ISBN 978-0671619466.
- Christiansen, Per; & Fariña, Richard A. (2004). Mass prediction in theropod dinosaurs. Historical Biology 16 (2-4): 85–92. doi:10.1080 / +08912960412331284313.
- Erickson, Gregory M.; Makovicky, Peter J .; Currie, Philip J.; Norell, Mark A .; Yerby, Scott A .; & Brochu, Christopher A. (2004). Gigantism and comparative life-history parameters of tyrannosaurid dinosaurs. Nature 430: 772–775. doi:10.1038 / nature02699.
- Therrien, François; & Henderson, Donald M. (2007). [http: //www.bioone.org/perlserv/? request = get-abstract & doi = 10.1671% 2F0272-4634 (2007) 27% 5B108% 3AMTIBTY% 5D2.0.CO% 3B2 & ct = 1 My theropod is bigger than yours ... or not: estimating body size from skull length in theropods]. Journal of Vertebrate Paleontology 27 (1): 108–115. doi:10.1671 / 0272-4634 (2007) 27 [108: MTIBTY] 2.0.CO; 2.
- Carr, Thomas D. (1999). «Craniofacial ontogeny in Tyrannosauridae (Dinosauria, Coelurosauria)». Journal of Vertebrate Paleontology 19 (3): 497—520
- Currie, Philip J. (2003). Cranial anatomy of tyrannosaurids from the Late Cretaceous of Alberta. Acta Palaeontologica Polonica 48 (2): 191–226. Архів [http: //app.pan.pl/acta48/app48-191.pdf оригіналу] за 14 квітня 2008. Процитовано 3 липня 2019.
- Holtz, Thomas R. (2004). «Tyrannosauroidea». in Weishampel, David B .; Dodson, Peter; & Osmólska, Halszka (eds.). The Dinosauria (Second ed.). Berkeley: University of California Press. pp. 111—136. ISBN 0-520-24209-2.
Посилання
- Дасплетозавр на Dinosaur.ru
- Discussion and specimen list at The Theropod Database. (англ.)
- Skull image of the Dinosaur Park Formation Daspletosaurus at The Graveyard. (англ.)
