Bombini
Bombini — триба, що належить до підродини Apinae та об'єднує сучасний рід Bombus та два вимерлих роди, а саме Calyptapis і Oligobombus.
? Bombini | ||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | ||||||||||||||||||||
| Біологічна класифікація | ||||||||||||||||||||
| ||||||||||||||||||||
| Bombini Latreille 1802 | ||||||||||||||||||||
Роди | ||||||||||||||||||||
|
Bombus
| ||||||||||||||||||||
Посилання | ||||||||||||||||||||
| ||||||||||||||||||||
Систематика
Традиційно у складі триби Bombini виділяють два самостійних роди — Bombus Latreille, 1802 та Psithyrus Lepeletier, 1832. Більшість сучасних (особливо іноземних) фахівців обидві групи з кладистичних міркувань об'єднують в один рід Bombus. Світова фауна роду Bombus налічує близько 250 видів, що відносяться до 15 підродів[1]. Зазначені підроди здається менш відрізняються один від одного, ніж підроди в більшості груп бджіл. Родові ознаки Bombus аналогічні ознакам триби.
Поширення
Джмелі є широко поширеною групою комах. Вони, як найбільш холодостійка група бджіл, здебільшого зустрічаються в холодному кліматі і найбільш поширені в Голоарктичній області, з більшою кількістю видів і підродів в Євразії, ніж у Північній Америці. На півночі ці комахи проникають до 82°30' в Канаді (о. Елсмір, Алерт) і 81°50' в Гренландії (мис Шмелке), а їх поширення обмежене наявністю ентомофільних рослин. На півдні вони поширені в Північній Африці і на захід аж до Канарських островів, але відсутні в Африці на південь від Сахари. На сході, види Bombus численні в Гімалаях та відмічені на висоті 5000 метрів, але відсутні нижче 1000 метрів над рівнем моря в Індії[2][3]. На південному сході кілька видів зустрічаються в горах Південно-Східної Азії (проте вони відсутні в низовинах) аж до островів Яви, Тайваню та Філіппінського архіпелагу. У західній півкулі є невелика фауна Bombus, ареал якої поширюється у всіх напрямках від архіпелагу Вогняної Землі, в основному в гірських районах або південно-помірних широтах, але на відміну від африканських і азійських видів, кілька видів Нового Світу поширені у вологих низовинних тропіках, таких, як долина річки Амазонки[3]. В даний момент вони відсутні тільки в Антарктиді і на деяких островах і архіпелагах. В Австралію, Нову Зеландію і окремі острова вони були інтродуковані для запилення культурних рослин[2].
В Україні зареєстровано 41 вид джмелів з яких на території лісостепової зони поширені 32 види, на території степової зони — 22, на території Полісся — 26, в Українських Карпат — 23, з яких 4 не трапляються ніде інде на території нашої країни, на території півострова Крим — 20[4].
Морфологія
Ейдономія
Форма голови з довгими щоками і збиральним апаратом на гомілках задніх ніг самок відразу вирізняють Bombini. За винятком паразитичних видів, триба представлена примітивними еусоціальними видами. Триба Bombini морфологічно одноманітна в порівнянні з Euglossini і, особливо, Meliponini. Тим не менш, спостерігається цікава різноманітність в кількох структурах, особливо в геніталіях самців, яйцекладі самок, і мандибулах. Велика частка таксономічних ознак групи ґрунтується на геніталіях самців. Серед Bombini зустрічаються середніх (довжиною 9 мм) та дуже великих (довжиною 22 мм) розмірів, волохаті бджоли, проте старі і частково безволосі особини іноді нагадують Anthophorini або Eucerini. Кігтики самок широко розставлені, аролії справжні але малі, на гомілках задніх ніг присутні шпори. За винятком соціальних паразитів, гомілка та перший членик лапки задніх ніг самки — засновниці та робочих особин подібні до робочих бджіл Apini. Джгутик також подібний до Apis. Крила мають повне жилкування. Радіальна комірка дещо довша, ніж відстань від її вершини до вершини крила. Птеростигма мала, загалом трохи довша ніж престигма, радіальна жилка виходить з радіальної комірки, що є прямою або ввігнутою. На задньому крилі відсутня югальна лопать. Восьмий метасомальний стерніт (S8) самця добре розвинений і склеротизований та має серединний відросток, що вкритий волосками. Диск стерніту не потовщений. Геніталії самців також склеротизовані з різноманітними гонобазами. Гонококсит великий і підтримує короткий і широкий гоностиль з ріноманітними утворами, що різняться у різних видів і широку, волохату вользеллу, що досягає вершини гоностиля, або перевищує його, а також виходить далеко у напрямку до основи гонококсита на нижній поверхні[3].
Мандибули самок на вершині зрізані або заокруглені. Нижній бік біля вершинного краю утворює кут біля вершини або зуб. Цей зуб іноді більш виражений і зубоподібний з невеликою виїмкою або розрізом в ньому над крайом. На зовнішній поверхні мандибул розташовані кілевидні гребені.
Личинка має невеликі загострені дорзолатеральні горбки на грудних сегментах. Мандибули дуже склеротизовані, їх вершини дещо зазубрені або плавно заокруглені, різко загострені в підроду Psithyrus, і мають невеликий передвершинний зуб на верхньому краю і велику верхівкову виїмку на внутрішній поверхні[3].
Органи чуття
Джмелі є тваринами-макросматиками, тобто мають сильно розвинений нюховий апарат. Сприйняття запахів здійснюється дистантними хеморецепторами, які у джмелів представлені різноманітними нюховими сенсили, розташованими на антенах.
Джмелі як слідові феромони використовують виділення мандибулярних залоз, такі, як транс-2,3-дигідрофарнезол і його ацетат, які застосовуються для розмітки території та приваблюють як самців, так і самок[5].
Імаго джмелів володіють двома типами органів зору: фасеткові або складні очі та дорсальні, або прості вічка.
В фасеткових очах джмелів наявні три типи фоторецепторів із специфічними опсинами, тобто трихроматична система кольорового зору, що є рідкістю серед комах. Наприклад, у джмеля Bombus distinguendus перший тип фоторецепторів має найбільшу чутливість до ультрафіолетового світла з довжиною хвилі 365 нм, пік чутливості другого типу фоторецепторів припадає на синій колір з довжиною хвилі 440—450 нм, а пік чутливості третього типу фоторецепторів припадає на зелений колір з довжиною хвилі 540—550 нм. Варто відзначити, що джмелі можуть бачити на квітках ультрафіолетові малюнки, приховані від погляду людини.
Для джмелів характерна найбільша швидкість сприйняття зорової інформації. В джмеля Bombus terrestris dalmatinus (його «зелені фоторецептори», що визначають ахроматичну швидкісну детекцію руху, працюють приблизно на третину швидше двох інших типів) швидкість сприйняття зорової інформації вдвічі більша за людську[6].
В Bombus на лобно-тім'яній поверхні голови розташовані три дорсальні вічка, що відповідають окремим фоторецепторам. Основна функція дорсальних вічок полягає в оцінці абсолютної освітленості та її змін.
Отруйний апарат та отрута джмелів
Отруйний апарат джмелів утворений кислою (з унікальною для триби розгалуженою будовою) та лужною залозами, що є видозміненими придатковими залозами яйцекладу і складно побудованим жалом, що є видозміненим яйцекладом та резервуара отруйного апарату. Таким чином отруйний апарат джмелів є прерогативою самок (фертильних і стерильних). Жало джмелів на відміну від медоносної бджоли позбавлене щербин, що дозволяє їм багаторазово жалити ворога[7].
.png.webp)
Отрута джмелів
При вжаленні джмелями спостерігається симптоматика, подібна до отруєння бджолиною отрутою: біль, набряк. Значна небезпека при поодиноких ужалениях не спостерігається, за винятком можливості розвитку алергічної реакції[8].
| Складник | Концентрація складника | Вид |
|---|---|---|
| Серотонін | Незначна кількість | Bombus terrestris |
| Ацетилхолін | 30 мг | Bombus terrestris |
| Фактор уповільнення м'язового скорочення (англ. slow muscle-contracting factor) | — | Bombus lapidarius |
| Путресцин | — | Bombus ignitus |
| MCD-пептид | 51 % сухої речовини | Bombus pennsylvanicus |
| Бомболітини | 25 % сухої речовини | Bombus pennsylvanicus |
| Цитрат | 6,5±8,2 % сухої речовини | Bombus fervidus, Bombus pennsylvanicus |
| Білки | 15±30 % сухої речовини | Bombus impatiens, Bombus pennsylvanicus |
В експерименті на кішках і щурах отрута земляного джмеля (Bombus terrestris) у дозі 100 мкг/кг при внутрішньовенному введенні викликає гіпотенгзивну реакцію, яка блокується атропіном і димедролом. У більш високих дозах (500 мкг/кг) отрута викликає порушення в діяльності серця. На перфузованому ізольованому серці жаби показано, що вона в концентраціях 10−8—10−7 г/мл володіє вираженою кардіотропною дією[10].
Путресцин
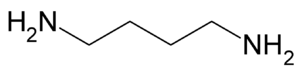
Цей біогенний амін (в хребетних) утворюється при гнитті білкових речовин під впливом декарбоксилаз деяких бактерій[11]. Летальна доза путресцину: для щура і кролика 1г/кг (підшкірно), для кролика 1,6 г/кг (перорально). Токсична дія путресцину проявляється в рухових парезах, судомах, уповільненні і аритмії пульсу, падіння кров'яного тиску і температури[12].
Путресцин є важливим компонентом рибосом та, ймовірно, залучений до утворення містків між різними ділянками цукровофосфатного остову рРНК[13].
Молекулярно-генетичні особливості Bombini
| Вид | Підрід | Диплоїдний набір | Гаплоїдний набір |
|---|---|---|---|
| Bombus nevadensis | Bombias | — | 17/18 |
| Bombus affinis | Bombus s.s. | — | 18 |
| Bombus moderatus | Bombus s.s. | — | 18 |
| Bombus occidentalis | Bombus s.s. | — | 18 |
| Bombus terricola | Bombus s.s. | — | 18 |
| Bombus griseocollis | Separatobombus | — | 18 |
| Bombus rufocinctus | Cullumanobombus | — | 18 |
| Bombus appositus | Subterraneobombus | — | 16 |
| Bombus borealis | Subterraneobombus | 32 | 16 |
| Bombus californicus | Fervidobombus | — | 19 |
| Bombus fervidus | Fervidobombus | 36 | 18 |
| Bombus pennsylvanicus | Fervidobombus | 36 | — |
| Bombus atratus | Fervidobombus | — | 20 |
| Bombus morio | Fervidobombus | — | 20 |
| Bombus perplexus | Pyrobombus | — | 12 |
| Bombus bimaculatus | Pyrobombus | — | 18 |
| Bombus impatiens | Pyrobombus | — | 18 |
| Bombus huntii | Pyrobombus | — | 18 |
| Bombus vagans | Pyrobombus | — | 18 |
| Bombus ephippiatus | Pyrobombus | — | 18 |
| Bombus melanopygus | Pyrobombus | — | 18 |
| Bombus mixtus | Pyrobombus | — | 18 |
| Bombus sitkensis | Pyrobombus | — | 18 |
| Bombus ternarius | Pyrobombus | — | 18 |
| Bombus vosnesenskii | Pyrobombus | — | 18 |
| Bombus ashtoni | Psithyrus | — | 25 |
| Bombus citrinus | Psithyrus | — | 26 |
Еколого-етологічі особливості джмелів в Україні
Еколого-етологічі особливості непаразитичних джмелів
В наших широтах джмелина сім'я існує один сезон. Восени матка, самці та робочі особини гинуть, а зимують лише молоді запліднені самки. Самки, що прокинулися після зимівлі мають слаборозвинені оварії, а тому певний час (до кількох тижнів) вони посилено живляться (за цей час рівень ювенільного гормону в їх гемолімфі підвищується та завершується розвиток оваріїв). У дозрілих самок проявляється гніздобудівельний інстинкт і вони починають шукати місце для гнізда[15].
Місце гніздування в джмелів є видоспецифічним — одні види будують гнізда в товщі ґрунту, інші на його поверхні або над ним. Деякі види, наприклад Bombus soroeensis (Fabricius,1777), характеризуються високою пластичністю щодо місця гніздування. При будуванні гнізда самки-засновниці не збирають гніздовий матеріал, використовуючи наявні в обраних місцях мох, сухі листки і стебла трав, деревна потерть, волосини тварин чи пух птахів, та не викопують ходи та порожнини в ґрунті, ця спільна риса для всіх представників родини Apidae відокремлює їх від споріднених антофорид (Anthophoridae), а натомість використовують покинуті нори дрібних ссавців, порожнини під коренями дерев, невеличкі ямки на поверхні ґрунту, трухляві пні, старі гнізда птахів, деревні дупла, шпаківні тощо У видів, які гніздяться на поверхні ґрунту, гніздо має форму напівсфери, тоді як форма підземних гнізд залежить від порожнини, в якій вони розміщені та зазвичай прикривають зверху гніздо спеціальним восковим куполом. Вхідний отвір самка-засновниця робить внизу однієї зі стінок, біля дна гнізда[2][15].
Посередині дна гнізда самка-засновниця формує першу виводкову камеру, а ближче до виходу восковий горщик, призначений для запасання меду. Розташувавшись на виводковій камері, головою до горщика, щоб надалі живитися з нього (запасів у горщику вистачає на добу), вона присупає до інкубації потомства за рахунок розігрівання власного тіла, завдяки швидкому скороченню крилових м'язів та інтенсивному обміну гемолімфи між середньогруддю і задньогруддю, і переривається лише під час вильоту для живлення. До появи робочих особин температура в гнізді коливається, адже одна самка-засновниця не здатна підтримувати її постійно. Наприклад, в гніздах B. terrestris діапазон коливання температури складає від 11 до 37 °C. Після вилуплення личинок, що відбувається за 3-5 днів з початку інкубації, самка-засновниця починає підгодовувати їх пилком та нектаром через тимчасовий отвір, який вона прогризає мандибулами в стінці виводкової камери. Закінчивши живлення, яке триває від 6 до 14 днів і залежить від умов навколишнього середовища, личинки плетуть кокони. Самка обгризає віск з коконів та споруджує з нього виводкову камеру[15].
Робочі особини в джмелів поділяються на фуражирів, які здобувають їжу, та внутрішньогніздових, які годують личинок, вентилюють та підтримують сталу температуру в гнізді, охороняють гніздо. Робочі особини перших виводків менші за матку, а їхні розміри зростають з ростом родини[15].
Нові камери будують поверх першої, вони мають вкрите воском дно. Форма та розташування камер в гнізді є видоспецифічною ознакою. Живлення личинок здійснюють робочі особини, при цьому їжу вводять через тимчасовий отвір в стінах виводкових камер, або виготовляють на стінках виводкових камер воскові кишені та наповнюють їх нектаром. За останнього типу вигодовування личинок стає можливою їхня конкуренція за їжу, яка створює варіабельність розмірів робочих особин в одному виводку. З ростом родини робочі особини розширюють гніздо. Після виведення кількох виводків робочих особин в родині з'являються репродуктивні особини[15].
Поява репродуктивних особин більшістю вчених пояснюється появою достатньої кількості корму для утримання останніх, що стає можливим за достатньої кількості робочих особин. Більш рання поява репродуктивних самок може бути викликана загибеллю матки. Робочі особини здатні відкладати яйця з яких виходять самці. Копуляція відбувається за 2-3 тижні після окрилення репродуктивних особин. Самки найчастіше копулюють одноразово[2][15].
Вороги непаразитичних джмелів
Гнізда непаразитичних джмелів, як і самі джмелі, піддаються нападу величезної кількості паразитів, інквілінів та хижаків[15].
Серед паризитів, що паразитують на Bombus потрібно відзначити нематоду Sphaerularia bombi (Dufour, 1837), мікроспоридію Nosema bombi (Fantham & Porter, 1914), личинки комах ряду двокрилих Conops flavipes (Linnaeus, 1758), Gonia fasciata (Meigen, 1826), Senotainia tricuspis (Meigen, 1838), Macronychia polyodon (Meigen, 1824) кліщі Parasitellus fucorum (De Geer, 1778), Tyrophagus laevis (Dujardin, 1849), Scutacarus acarorum (Goeze, 1780), Locustacarus buchneri (Stammer, 1951), Acarapis woodi (Rennie, 1921), браконіду Syntretus splendidus (Marshall, 1887), патогенні грибки Beauveria bassiana ((Balsamo-Crivelli) Vuillemin, 1912), Paecilomyces fumosoroseus (Wize) A.H.S.Br. & G.Sm., (1957), Cephalosporium lecanii (Zimm., 1898), Aspergillus candidus (Link, 1809). Личинки жуків-майок (Meloe), жуків-пістряків (Cleridae), мух-тахін (Tachinidae) живляться розплодом джмелів. Личинками та лялечками джмелів живляться личинки ос-німок (Mutillidae) і хальцид (Chalcidoidea)[15].
Схильність до клептопаразитизму характерна для багатьох підродів та видів джмелів, що проявляється за несприятливих умов. Облігатними інквілінами серед джмелів є підрід джмелі-зозулі (Psithyrus), що паразитують на інших підродах джмелів. В обох випадках паразит та хазяїн морфологічно і фізіологічно подібні[15].
На імаго джмелів полюють ктирі (Asilidae) та бджолині вовки (Philanthus triangulum (Fabricius, 1775)), павуки-краби (Thomisidae) підстерігаючи останніх на квітках. Досить часто робочі особини джмелів потрапляють в ловчі сітки павуків. Джмелі є складовою частиною раціону бджолоїдки, осоїда, великої синиці (її жертвами найчастіше стають джмелі зі взятком, яким вона живиться), ворони, сорокопуда та інших птахів. Мурахи, сірі полівки, миші (хатня, польова, лісова та ін.), кроти, їжаки, борсуки, лисиці розорюють гнізда джмелів[15].
Медоносні бджоли (Apis mellifera Linnaeus,1758) можуть нападати на гнізда джмелів та викрадати мед[15].
Інфекційні та паразитарні хвороби Bombini
| Інфекційні та паразитарні хвороби Bombini [16], [17] | ||||||
| Патоген | Хвороба | Вид | Стадія розвитку | Ознаки та симптоми | ||
|---|---|---|---|---|---|---|
| Віруси | ||||||
| Вірус гострого паралічу бджіл (англ. Acute bee paralysis virus, ABPV) | Гострий параліч бджіл | Bombus terrestris | Імаго | Імаго можуть бути паралізованими або симптоми можуть бути відсутні. Вважається, що не передається орально, але виявляється в обніжжі фуражирів та торакальних слинних залозах. Може бути перенесений Varroa destructor. | ||
| Вірус чорних маточників (англ. Black queen cell virus, BQCV) | Чорний маточник | Bombus spp. | Личинка самки-засновниці, Імаго | — | ||
| Вірус деформованих крил (англ. Deformed wing virus, DWV) | Деформованість крил | Bombus spp. | Імаго | — | ||
| Вірус бджіл Кашміру (англ. Kashmir bee virus, KBV) | Хвороба Кашміру | Bombus terrestris | Імаго | Вірус бджіл Кашміру та Ізраїльський вірус гострого паралічу бджіл (англ. Israeli acute paralysis virus, IAPV) тісно пов'язані між собою, і найкраще описані для медоносних бджіл, всі стадії розвитку яких можуть інфікуватися. Передача: горизонтальна від заражених дорослих через залозисті виділення, що контактують з джерелами живлення; трансоваріальна; і передається вектором Varroa destructor. Інфікований розплід або гине до того, як стільник закривається, або відновлюється після інфекцій. Інфекції можуть зберігатися у дорослих без симптомів або викликати швидку смертність протягом декількох днів. Віріони безоболонкові (прості), ікосаедричний капсид, діаметром ~ 30 нм. | ||
| Вірус мішкуватого розплоду (англ. Sacbrood virus, SBV) | Мішкуватий розплід | Bombus spp. | Личинка, Імаго | — | ||
| Мікроспоридії | ||||||
| Nosema bombi | Нозематоз | Bombus terrestris, Bombus occidentalis, Bombus impatiens | Імаго | Хвороба характеризується наявністю спор у всій порожнині тіла (~ 4.2-5.4 × 2.7-3.5 мкм) з нечисленними чіткими позначками. В Bombus terrestris і Bombus impatiens, ріст колонії та продуктивність репродуктивності значно знижуються, але не в Bombus occidentalis.
Передача трансоваріальна й оральна. | ||
| Протисти | ||||||
| Apicystis bombi | — | Bombus affinis, Bombus bimaculatus, Bombus fervidus, Bombus griseocollis, Bombus impatiens, Bombus perplexus, Bombus terricola, Bombus vagans | Імаго | Поганий ріст колонії, жирове тіло знищено у заражених робочих особин. Великі спори, виявлені в фекаліях. | ||
| Crithidia bombi та C. expoeki | — | Bombus terrestris | Імаго | Трипаносоми, які трапляються у задній кишці джмелів. Передача відбувається горизонтально всередині гнізда, між гніздами через відвідування квітів та трансоваріально. Інфекції впливають на засновницьку здатність нових маток, виживання робочих особин та відтворення колоній. | ||
| Кліщі | ||||||
| Acarapis woodi | — | Bombus spp. | Імаго | В уражених джмелів можуть бути розірвані крила і труднощі при польоті, роздуте черевце, або симптоми можуть бути відсутні. Кліщі трапляються в проторакальній трахеї. Самки мають довжину 143-174 мкм, самці 125-136 мкм, з дещо прозорою, гладкою кутикулою з декількома довгими волосками на тілі й ногах. Вплив на розмір і продуктивність колонії є невизначеним. | ||
Примітки
- Williams P.H., Cameron S.A., Hines H.M., Cederberg B., Rasmont P. A simplified subgeneric classification of the bumblebees (genus Bombus) // Apidologie. — 2008.-Vol. 38.-P. 1- 29
- Радченко В. Г., Песенко Ю. А. Биология пчел (Hymenoptera, Apoidea). СПб.: Зоол. ин-т РАН,1994.- 450 с.
- Michener, C. D. The Bees of the World. — Baltimore: Johns Hopkins University Press, 2007. — 913 p.
- Konovalova I. B. The Bumble Bees of Ukraine: Species Distribution and Floral Preferences // Psyche: A Journal of Entomology.— 2010.— Vol. 2010 (2009).—P. 44-54.
- ФЕРОМОНЫ НАСЕКОМЫХ (ИВАНОВ В. Д., 1998), БИОЛОГИЯ
- Differences in Photoreceptor Processing Speed for Chromatic and Achromatic Vision in the Bumblebee, Bombus terrestris
- Гелашвили Д. Б., Крылов В. Н., Романова Е. Б. Зоотоксинология: биоэкологические и биомедицинские аспекты. Учебное пособие: Нижний Новгород: Изд-во ННГУ, 2015. — 770 с.
- Орлов Б. Н. и др. Ядовитые животные и растения СССР: Справочное пособие для студентов вузов по спец. «Биология»/Б. Н. Орлов, Д. Б. Гелашвили, А. К. Ибрагимов. — М.: Высш. шк., 1990. — 272 с.
- Bucher C, Korner P, Wuthrich B. Allergy to bumblebee venom. Curr Opin Allergy Clin Immunol 2001;1(4):361-5.
- Орлов Б. Н., Гелашвили Д. Б. Зоотоксинология (ядовитые животные и их яды) — М.: Высш. шк., 1985. — 280 c.
- Рево, А. Я. Органическая химия: для студ. мед. ин-тов. — М.: Высшая школа, 1962. — 383 с.
- Краткая химическая энциклопедия. — М.: Советская энциклопедия. — 1965. — Т. 4. — 1105 c.
- Спирин А.С. Молекулярная биология. Рибосомы и биосинтез белка. — М.: Академия, 2011. — 513 с.
- R. E. Owen, K. W. Richards and A. Wilkes. Chromosome Numbers and Karyotypic Variation in Bumble Bees (Hymenoptera: Apidae; Bombini). Journal of the Kansas Entomological Society, Vol. 68, No. 3 (Jul., 1995), pp. 290-302.
- Горобчишин В. А., Подобайло А. В. Біологічні особливості та охорона джмелів: Навчальний посібник. — Київ: Геопринт, 2004. — 32с.
- Vega F. E., Kaya H. K. Insect Pathology. — San Diego: Academic Press, 2012. — 508 p. — ISBN 978-0-12-384984-7. (англ.)
- Руководство по вирусологии: Вирусы и вирусные инфекции человека и животных / Под ред. академика РАН Д.К. Львова. — М.: ООО «Издательство «Медицинское информационное агентство», 2013. — 1200 с. — ISBN 978-5-9986-0145-3. (рос.)