Гаплогрупа R1b
Гаплогру́па R1b (R-M343; Hg1; Eu18; R-L21) — гаплогрупа людської Y-хромосоми. Разом із гаплогрупою R1a є відгалуженням гаплогрупи R1. Ймовірно, походить із Західної Азії[1]. Найбільшої частоти серед чоловічого населення сягає у країнах Західної Європи (Уельс, Шотландія, Ірландія — 90 %, країна басків — 88 %, Іспанія — 70 %, Франція — до 60 %[2], Португалія — понад 50 %[3]). Також поширена серед деяких племен Чаду, серед кількох племен туркмен та башкир, іранських ассирійців (55 %[4]), російських кумандинців (до 49 %)[5]. З меншою частотою поширена у Центральній та Північній Європі (Західна Німеччина — 47 %, Італія — 40 %[6], Норвегія — близько 30 %[7]), на Балканах, в Малій, Західній та Центральній Азії, у Північній Африці, серед мовців алґських мов у Канаді. За даними Familytree DNA[8][9][10], 35 з 601 (5,8 %) чоловіків-українців є носіями R1b.
.png.webp)
Чисельні представники цієї гаплогрупи у популяції Америки та Австронезії, що утворилися через хвилі переселенців з Європи у новітній час. Щодо високого рівня у доколумбових народів-мовців алгських мов у Східній Канаді від древньої міграції тривають дискусії серед науковців. Вживаються інші позначення гаплогрупи — Hg1 , Eu18.
Історія дослідження
Три аутосомних дослідження у 2015 році підтвердили курганну теорію Марії Гімбутас про батьківщину індоєвропейців. Відповідно до цих досліджень гаплогрупи R1b (найпоширеніша у Західній Європі) та R1a (найпоширеніша у Східній Європі та Південній Азії) поширилися разом з індоєвропейськими мовами з українського та російського степу. Ці дослідження також вказали на аутосомний компонент у сучасних європейцях, що не був присутній у європейцях новокам'яної доби й які з'явилися разом з батьківськими лініями R1b й R1a, та індоєвропейськими мовами.[11][12][13]
Місце появи R1b лежить у Євразії й найімовірніше у Західній Азії.[14] Т. Карафет оцінив у 2008 році вік батьківської для R1b гаплогрупи R1 у 18500 років тому.[15]
Раніше вважалося, що гаплогрупа R1b походить з Європи. Так у 2000 році Орнелла Семіно з колегами аргументували, що R1b була ще до кінця льодовикової доби й з відступом льодовика поширилася на північ з притулку на Іберійському півострові.[16] Оцінки часу перебування щонайменше до більшості R1b у Європі поступово зменшувалися для новокам'яної доби або пізніше.[14][17][18][19] Проте у 2010 році аргументував за давньокам'яне походження R1b1b2 у Європі.[20] Чіхі вказав, що час генетичних розходжень на який вказують генетичні підрахунки не збігається з розділеннями населення. Тобто час життя найближчого загального предка гаплогрупи R1b, що жив у Євразії (можливо у давньокам'яну або новокам'яну добу) не вказує на заселення ним Західної Європи.[21] Підсумовуючи, Майкл Р. Магліо аргументує що найближча гілка R1b є з Іберійського півострову та його субклади у Західній Азії та у Африці є прикладом задньої міграції не з її місця походження.[22] Отже Барбара Арреді вперше вказала на поширення мікросателітних варіацій скотарів гаплогрупи R1b у Європу зі східних степів на захід, коли вхід у Європу зі Західної Азії більше відноситься до поширення сільськогосподарських племен.[19] У 2009 році Чіароні додав до цього погляду, що поширення гаплогрупи R1b є показовим для хвильового поширення гаплогрупи, у даному випадку це стосувалося поширенню зі сходу на захід.[23] Пропозиція про прихід у Європу R1b з південного сходу базувалося на 3-х детальних дослідженнях великих баз даних 2010 року. Вони вказали на найраніші субклади R1b знайдені у Західній Азії та найпізніші — у Західній Європі.[14][17][24]
Зовнішня структура
Гаплогрупа R1b є частиною гаплогрупи R (M207). Її попередницею є гаплогрупа P1 (M45), що є частиною гаплогрупи P (P295). У межах гаплогрупи K2b (P331) вона споріднена за походженням з гаплогрупами M, P, Q, R, S. Гаплогрупа K2 (M526) відноситься до макро-гаплогрупи K (M9), що згрупувала майже всіх представників чоловічої статі поза Африкою.
- IJK
- IJ (M429/P125)
- I (M170)
- J (M304)
- K (M9)
- IJ (M429/P125)
Внутрішня структура
- R1b (M343)
- R1b* (M343*) — надто нечисельні, переважно у Ірані;
- L278 (M415) — найчисельніші
- L278* (M415*) — початкові форми L278 (M415) виявлені у людей самарської культури мідної доби. Сучасні на українському Передкарпатті, менше у Східній, Західній Азії, в Італії
- PH155 — надзвичайно рідкісне виявлені у Східному Туркестані, Таджикистані, Бутані, на Бахрейні;
- M335 — з'явилася 7400 років тому; надзвичайно рідкісна виявлена у Туреччині;
- PH200 — з'явилася 7400 років тому;
- Y32792 — з'явилася 5800 років тому;
- Y81807 — з'явилася 5800 років тому;
- L754 — найчисельніші
- L754* — віллабрунська людина епіграветської культури з північної Італії; сучасний народ півдня Сахари сахарської мовної сім'ї;
- V2219 — з'явилася 17100 років тому;
- L389
- V1636 — дуже рідкісні у різних частинах південно-західної Азії, на Середземномор'ї та у континентальній Європі
- P297 — більшість євразійських R1b належить цьому субкладу, представляючи велику частину сучасного людства.
- P297* — виявлена у ямній культурі бронзової доби
- M478 — нащадки половців-кипчаків серед казахів, башкир і козаків; серед балкарців та чуваш (нащадки булгар); гіпотетичні залишки тохарів і саків індоєвропейомовного Турану;
- Y14051
- Y20750
- M269 — найчисельніша гаплогрупа R1b, що складає більшість чоловічого населення Західної Європи; поширена в Євразії від понад 90 % у Ірландії та Велсі до 1 % у Індії та Китаї; у Африці з максимумом в Алжирі.
- RF7562
- Y36978
- RF7563
- L23 — є найбільш поширеними з M269
- L23* — зустрічається у Анатолії, на Кавказі та Середземномор'ї.
- Z2103
- RF331
- Y4364
- L584
- Z2106
- L51
- L51* — зустрічається з найбільшою частотою у південній Франції та північній Італії.
- Z2118
- Z2116
- Y5141
- S1161
- Z2116
- L151 - включає більшість чоловіків з R1b у Західній Європі;
- A8039
- A8051
- S1200
- FGC30867
- Y36365
- S17624
- S14328
- U106 - до неї належать 25 % носіїв R1b у Європі з центром поширення у Фрісландії й у Центральній Європі;
- S16185
- S18632
- Y3443
- FGC3861
- Z19
- S263
- P312
- Y18209
- PH2278
- DF99
- DF19
- L238
- U152
- BY3851
- Y22161
- A275
- Y22447
- CTS2617
- Z192
- Z56
- Z36
- L2
- DF27
- S46
- L21
- RF7562
R1b* (M343*)
M343 сформувалася 22800 років тому. Загальний предок жив 20400 років тому.
Чоловіки з гаплогрупи R1b* без подальших мутацій надто рідкісні. Вони складають 13 % серед курдів південно-східного Казахстану, 4,3 % серед персів провінції Єзд у центральному Ірані й 3,2 % серед азербайджанців іранського Азербайджану[25]
R1b1 (L278 або M415)
L278 або M415 сформувався 20400 років тому. Загальний предок жив 18900 років тому.
Представники початкової форми L278 (M415) та PH155 рідкісні серед сучасних чоловіків. Проте до неї належать древні 2 скелети енеолітичної самарської культури з північного Надкаспію приблизно 5600 років до Р. Х. та чоловік ранньої новокам'яної кардіальної кераміки культури з Елс-Трокс у Піренеях Арагону датований приблизно 5100 роками до Р. Х..
Серед сучасних зразків у 2010 році Варзан виявив 3 випадки з 322 (0,93 %) у карпатсько-дністровському краї України. Інші популяції M415: східні азіати — 1 з 156 (0,64 %) ; західні азіати — 1 з 328 осіб (0,3 %); італійці — 3 випадки з 1173 (0,26 %).[26]
Представники відгалужень L278 є найчисельнішими R1b.
Найближче розгалуження дерева R-L278 або R-M415:
- L278 (M415)
- M415*
- L754 — з'явилася 20400 років тому;
- L389 — з'явилася 17100 років тому;
- P297 — з'явилася 15600 років тому;
- V1636 — з'явилася 15600 років тому;
- V2219 — з'явилася 17100 років тому;
- V88 — з'явилася 15000 років тому;
- L389 — з'явилася 17100 років тому;
- PH155 — з'явилася 20400 років тому;
- M335 — з'явилася 7400 років тому;
- PH200 — з'явилася 7400 років тому;
- Y32792 — з'явилася 5800 років тому;
- Y81807 — з'явилася 5800 років тому;
R1b1a (L754)
L754 сформувалася 20400 років тому. Загальний предок жив 17100 років тому.
L754 мала давня людина з Віллабруна-1 епіграветської культури, що жила 14000 років тому у північно-східній Італії.
L761 (еквівалент для L754) має високе поширення (34 %) серед сахарсько-мовного народу Тубу у Чаді.
R1b1a1 (L389)
L389 сформувалася 17100 років тому. Загальний предок жив 15600 років тому.
L388/PF6468 та L389/PF6531 у їх первісній формі дуже рідкісні або зникли. Її субклади відносно рідко зустрічаються у різних частинах південно-західної Азії, на Середземномор'ї та у континентальній Європі.
R1b1a1a (P297)
P297 сформувалася 15600 років тому. Загальний предок жив 13300 років тому.
P297 є предком для значних субкладів M73 та M269, що об'єднує їх у одне гроно.
Скелет ямної культури мідної доби у районі Самарської Луки датується 3305–2925 років до Р. Х. є R1b1a* тестований позитивно на маркер P297, але негативний для L51.
Більшість євразійських R1b належить цьому субкладу, представляючи значну частину сучасного людства.
Відгалуження P297 незрівнянно добре вивченні порівнюючи з самим початковим субкладом P297.
Найближче розгалуження дерева R-L297:
- P297
- M269
- L23
- RF7562
- M478 та M73
- Y14051
- Y20750
- M269
Східні, степові M73 і M478
R1b1a1a1a (M478) сформувалася 13300 років тому. Загальний предок жив 7300 років тому.
R1b1a1a1 (M73) та R1b1a1a1a (M478) обминули Каспій з Півночі й заселили Туран. За іншою версією потрапили у Туран через сучасну території Азербайджану, Ірану та Туркменістану. Згодом були відомі як саки. Носії R-M478 були носіями давніх індоєвропейських мов Центральної Азії й тюркською експансією стали носіями переважно тюркських мов.
Домінантна гаплогрупа серед алтайського народу кумандинців, що мають походження від куманів (половці). Також гаплогрупа поширена серед етнічної групи кипчаків, що складають 6-7 % казахського народу; серед кипсаків у складі башкир (у південно-східних районах до 23,4 %); серед нащадків запорозьких козаків у складі українського народу. Казахські кипчаки та башкирські кипсаки походять від історичних кипчаків-половців. За Походження тюркомовних кипчаків пов'язується з іраномовними саками Турану.[27][28]
Раніше за дослідженням Сенгупта 2006 року були помилково ідентифіковані як належні R-M73 представники хазарейців Афганістану, що за новітнім дослідженням Бехара 2010 року належать до гаплогрупи R2 та Q.
Подібно багато башкир, що раніше відносили до M478 виявилися представниками U-152 (R1b1a1a2a1a2b) та деякі, переважно з південного сходу Башкортостану — представниками гаплогрупи Q-M25 (Q1a1b).
Найближче розгалуження дерева R-M478:
- M478 — з'явилася 13300 років тому;
- Y20750 — з'явилася 7200 років тому;
- Y91288 — з'явилася 5700 років тому; поширена серед балкар Кабардино-Балкарії;
- Y20748 — з'явилася 5700 років тому; поширена серед татар у Татарстані;
- Y14051 — з'явилася 7200 років тому; поширена серед нащадків українських козаків в Україні й Росії;
- Y134928 — з'явилася 1550 років тому;
- Y53944 — з'явилася 1550 років тому; поширена у Росії серед нащадків половців Надволжя (Саратовська й Нижегородська область).
- Y20750 — з'явилася 7200 років тому;
R1b1a1a2 (M269)
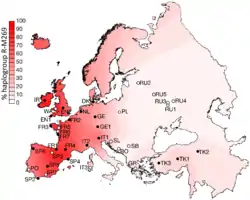
M269 сформувалася 13300 років тому. Загальний предок жив 6400 років тому.
Гаплогрупа R-M269 щільно поширена у Європі.
На 2017 років R-M269 або R1b1a1a2 є найпоширенішою Y-ДНК європейських чоловіків. Носіями є приблизно 110 мільйонів європейців.
R-M269 є однією з самих досліджених Y-ДНК гаплогруп у плані поширення індоєвропейців до Європи. Особливо R-L23 (R-Z2103) була знайдена превалентною у стародавніх зразках що відносяться до ямної культури.
Батьківщиною M269 є Західна Азія. З'явилася у Європі у новокам'яну добу. Проте різноманіття субкладів M269 у Європі та їх географічне поширення пов'язані з колонізаційними хвилями з євразійських степів за мідної та бронзової діб.
Поширення M269 зростає зі сходу на захід Європи. Вона досягає максимуму 92 % у Велсі, 82 % в Ірландії (у деяких місцях до 95 %), 70 % у Шотландії, 68 % в Іспанії, 60 % у Франції (76 % у Нормандії), близько 60 % у Португалії, 53 % в Італії, 50 % у Німеччині, 50 % у Нідерляндах, 45 % у східній Англії, 43 % у Данії та 42 % в Ісландії. Мінімально, у Західній Європі може сягати, наприклад 27 % у верхів'ях Рони. З меншою частотою M269 зустрічаються у Центральній (наприклад 2,4 % у Польщі) та Південно-Східній Європі та зменшується Євразією до менше 1 % у Китаї та Індії. У Африці найбільшого поширення досягає у Алжирі (10 %) та серед гереро у Намібії (8 %).
Західноєвропейці M269 поділені на представників P312/S116 та U106/S21 відгалуження M412 (L51).
Поза оригінальними початковим R-M269 решта ділиться на 2 головні групи:
- R-L23 (R1b1a1a2a; L23/PF6534/S141) — сформована 6400 років тому: загальний предок 6100 років тому.
- R-PF7562 (R1b1a1a2b; V2850) — сформована 6400 років тому: загальний предок 5600 років тому.
R-L23* є найбільш поширеними R-M269. коли R-PF7562 тепер рідкі.
У Росії R-L23 (Z2105/Z2103; або R1b1a1a2a) за дослідженням 2015 року виявлена у 36,2 % башкир Бурзянського району, 21,2 % удмуртів, 8 % комі, 6,8 % мордовців, 3,8 % бесерм'ян та 2,3 % чуваш.
На Балканах R-L23 виявлена у 7,9 % косівських албанців, 5,1 % македонців, 4,4 % сербів.
Найближче розгалуження дерева R-M269:
- M269
- L23
- L51
- Z2103
- RF7562
- RF7563
- Y36978
- L23
R1b1a1b (R-V1636)
V1636 на сьогоднішній день немає субкладів. Він є дуже рідкісним і зустрічається у Середземномор'ї, й особливо у Болгарії.
R1b1a2 (R-V88)
V88 сформувалася 17100 років тому. Загальний предок жив 11800 років тому.
Гаплогрупа, що визначається сніпом V88 була виділена Крусіані у 2010 році. Носіями її є народи півдня Сахари афроазійської мовної сім'ї її чадської гілки.
у їх первісній формі дуже рідкісні або зникли. Її субклади відносно рідко зустрічаються у різних частинах південно-західної Азії, на Середземномор'ї та у континентальній Європі.
Галузки V88 найбільше поширена серед у північному Камеруні на заході центральної Африки:
- берберів (26,9 % берберів з Сіви у Єгипті, 4,5 % туарегів Нігеру, 3 % мозабільські бербери Алжира),
- семітів (40,0 % шува араби у Камеруні; 5,8 % південних єгиптян, 4,9 % бахарія у Єгипту; 4,1 % північних єгиптян 0,3 % арабів Марокко),
- чадські народи (95,5 % оулдеме у Камеруні; 87,5 % мафа у Камеруні; 82,4 % мада у Камеруні; 77,8 % гуізіга у Камеруні; 66,7 % гуїдар у Камеруні; 42,1 % даба у Камеруні; 28,6 % масса у Камеруні; 20,0 % хауса на півночі Нігерії),
- західноатлантичних народів (14,3 % фульбе у Нігері, 11,1 % фульбе у Камеруні),
- ніло-сахарських народів (14,3 % канурі у Камеруні; 9,1 % нгамбай у Чаді),
- бенуе-конголезькі народи (4,8 % йоруба у Південній Нігерії),
- адамауа-східні народи (66,7 % моунданг у Камеруні; 20,8 % фалі у Камеруні; 9,1 % талі у Камеруні).
Високі відсотки на заході центральної Африки обумовлене до-ісламським рухом народів з Євразії.
Найближче розгалуження дерева R-V88:
R1b1b (PH155)
PH155 сформувалася 18900 років тому. Загальний предок жив 7300 років тому.
R-PH155 (R1b1b) є надзвичайно рідкісне й визначається присутністю сніпа PH155.[30] ISOGG на 2017 рік визначає дві галузі R-M335 (R1b1b1) та R-PH200 (R1b1b2).
Були виявлені у Східному Туркестані, Таджикистані, Бутані, на Бахрейні.
M335 уперше було задокументоване у 2004 році, коли було виявлене у Туреччині.
R1b серед українців
За даними трьох груп Familytree DNA[8][9][10], де сконцентровані українці, носіями R1b є 35 з 602, тобто 5,8 %.
- Усі носії R1b відносяться до Р297.
- до «європейської гілки» М269 відносяться 31 з 35 (88,6 %) R1b; взагалі серед українців М269 — 5,15 %. Усі визначенні за галуззю (числом 17) українці-носії М269 відносилися до L23. З них до:
- «західноєвропейська» L51 та нижче за деревом L151 відносяться 11 з 17 (64,7 %). Носії L151 розділені на:
- «германська» U106 (7 з 11). До U106 відноситься:
- 4 зразка L48 (у тому числі 2 до Z331)
- 1 зразок до S264,
- P312 (4 з 11). До P312 відноситься:
- 2 зразка L2,
- 1 зразок DF27,
- «германська» U106 (7 з 11). До U106 відноситься:
- «балкано-малоазійська» Z2103 — 5 з 17 (29,4 %). Серед носіїв Z2103:
- 4 зразка належить до Z2108 та вниз за деревом до Z2110
- 3 зразхка CTS7556,
- 1 зразок CTS7822,
- 1 зразок належить до Z2109,
- 4 зразка належить до Z2108 та вниз за деревом до Z2110
- «західноєвропейська» L51 та нижче за деревом L151 відносяться 11 з 17 (64,7 %). Носії L151 розділені на:
- до «половецько-козацької гілки» М478 — 4 з 35 (11,4 %); взагалі серед українців М478 — 0,66 %.
- до «європейської гілки» М269 відносяться 31 з 35 (88,6 %) R1b; взагалі серед українців М269 — 5,15 %. Усі визначенні за галуззю (числом 17) українці-носії М269 відносилися до L23. З них до:
Історія походження
Гаплогрупа R1b відноситься до макро-гаплогрупи K (M9), що згуртувала майже усіх представників чоловічої статі поза Африкою. Її батьківщиною виникнення є Азія.
Скотарі північної Месопотамії
Мисливські племена R1b перетворилися на скотарів. Після того як мамути зникли у степах Північної Месопотамії мисливці перейшли на полювання бізонів та турів. Коли населення Месопотамії збільшилося приблизно 12000 років тому популяції турів, кабанів, кіз та полювання на них стало регульованим.
За генетичними дослідженнями сучасні Bos taurus походять від популяції 80 турів. Найранні свідчення про одомашнення худоби виявлено у докерамічній новокам'яній археологічній культурі у Таврійських горах. Розташовані на відстані у 250 км два поселення новокам'яної доби Чайоню-Тепесі (південно-східна Туреччина) та Джа'де ель-Мугхара (північний Ірак) 10500 років тому мали одомашненили корів Bos taurus. Саме цей край був батьківщиною для мисливців R1b де вони перетворилися на скотарів. Вважається що у одомашненні брали учаслть плнмена J2. Найбільше різноманіття архаічних типів R1b зустрічається у Східній Туреччині (Анатолії). Саме звідси більшість дослідників виводять місце походження гаплогрупи.[31]
Саме з цієї місцевості північної Месопотамії поширилися три головні гілки R1b:
- R1b1a (P297) — північна група виникла у північній Месопотамії, східній Туреччині, Закавказзі, та згодом перетнула Кавказькі гори у Понтійсько-каспійський степ з ідеальними пасовиськами для корів. Тут племено P297 розділилося на дві галузі:
- R1b1a1 (M478) — обминули Каспій з Півночі й заселили Туран. За іншою версією потрапили у Туран через сучасну території Азербайджану, Ірану та Туркменістану. Згодом були відомі як саки. Носії R-M478 були носіями давніх індоєвропейських мов Центральної Азії й тюрською експансією стали носіями переважно тюркських мов;
- R1b1a2 (M269) — найбільш чисельна гаплогрупа R1b деякий час концетрувалася у степах між Волгою та Дунаєм (ямна культура) й згодом заселили через шлях Дунаєм Західну Європу, де склали більшість сучасного чоловічого населення. Сучасні носії R-M269 стали носіями індоєвропейських мов;
- R1b1b(M335) — анатолійська група, що зустрічається в Туреччині. Найбільш неуспішна група R1b, що залишилася малочисельною за недостатком пасовиськ в Анатолії;
- R1b1c (V88), — південна галузь найчисельна на Леванті та на південь від центральної Сахари у Африці. Мігрувала у південно-західному напрямі у Єгипет й згодом освоїли усю Північну Африку з простіром Сахари, що тоді була суцільним степом.
R-P297 у понтійсько-каспійському степу
Племена P297 населяли Понтійсько-каспійський степ й були носіями курганної культури у 4200-2200 роках до Р. Х. Кінь був одомашнений приблизно 4600 років до Р. Х. у степах між Доном та Волгою попередниками R-P297 племенами R1a або найстародавнішими галузями P297 (хвалинська культура). Ранні носії P297 аутосомно іноді мають додаток сибірської домішки та відсутність кавказької домішки. Пізні археологічні культури мідної доби, як афанасьївська, ямна та шнурової кераміки усі мають аутосомну кавказьку домішку.
Скелет ямної культури мідної доби у районі Самарської Луки датується 3305–2925 років до Р. Х. є R1b1a* тестований позитивно на маркер P297, але негативний для L51.
Доба дніпро-донецької культури
R-M269 є «європейською» галуззю гаплогрупи R-P297. Достеменно невідомо коли R-M269 перетнула Кавказ у понтійсько-каспійський степ, але припускають що то було з появою дніпро-донецької культури у 5100-4300 роках до Р. Х.. Дніпро-донецька культура була першою спражньою новокам'яною спільнотою у цих степах з одомашненими коровами, вівцями та козами. За доби дніпро-донецької культури відбувається перехід від племен R1a або/та I2a1b до раннього землеробства, можливо з домішкою близькосхідних фермерів «Старої Європи».
Понад 30 зразків з України новокам'яної доби (5500-4800 роки до Р. Х.) були досліджені Матієсоном[32] у 2017 році. Вони належать до Y-гаплогрупи I, I2a2, R1a, R1b1a (L754) та лише один до R1b1a2 (L388). Жодний не належить до R1b-M269 або R1b-L23, які стали домінуючими у добу ямної культури.
Мітохонріальні результути належать U4a, U4b, U4d, U5a1, U5a2, U5b2, а також по одному до J2b1 та U2e1. Жодна з цих мітохондріальних гаплогруп не належить до типових індо-європейських H2a1, H6, H8, H15, I1a1, J1b1a, W3, W4 або W5 що з'являються пізніше за ямної, шнурової кераміки та унетицької культур. Аутосомні геноми новокам'яної України були чисто з середньокам'яної доби Європи (приблизно 90 % EHG та 10 % WHG) та повністю відсутні кавказькі домішки (CHG), які вже знайдені за часу ямної культури та культур-наступниць бронзової доби.
Доба середньостогівсько-хвалинської культурно-історичної спільності
Перша чисто прото-індоєвропейською культурою була середньостогівсько (4600-3900 роки до Р.Х.)-хвалинська (5200-4500 роки до Р.Х.) культурно-історична спільнота. Саме тоді з'являються та продовжуються у наступних степових культурах самобутні поховання під могилами (курганами) з положенням померлих на спині з піднятими, зігнутими колінами, з напрямом голови на північний схід.
Доба майкопської культури
Інша міграція з Закавказзя на Передкавказзя сталося 3700 років до Р. Х. з появою майкопської культури бронзової доби. Майкопська культура з'явилася на Кубані з переходом племен мідної доби Асирії та Мідії через Кавказ. Майкопська культура поширила бронзову технологію та обряд поховання у понтійсько-каспійський степ, де, незабаром (з 3500 року до Р.Х), сформувалася ямна культура.
Майкопська культура (3700-2500 роки до Р. Х.) була південною частиною ямної культурно-історичної спільноти.
Попередниця майкопської культури — свободненська культура (400-3700 роки до Р. Х.) мала зв'язки з суворівською групою скелянської культури та новоданилівською культурою. Кемі-обинська культура Криму та Півдня України утворилася міграцією Майкопської культури у середовище нижньомихайлівської та середньостогівської культур. Навіть старший (5000-4500 роки до Р. Х.) подібний до хвалинської культури Нальчинське поселення у Кабардії вказує на культурно-племінну міграцію з Передкавказзя у понто-каспійські степи. Саме у цей період R1b почали відносини й змішування з їх попередниками у понто-каспійських степах R1a.
Майкопська та ямна культури мали однаковий обряд поховання — померлих ховали у всипаних охрою ямах горілиць з піднятими у гору зігнутими колінами з головами у напрямі північного-сходу, тобто сходу сонця. У поховальних ямах були жертовна домашня худоба. Вони користувалися кіньми, критими возами, з переважно коров'ячим стадом і меншим числом вівці для шерсти. Також користувалися мідними та бронзовими виробами.
Проте деки елементи майкопської культури докорінно відрізняються, що вказує на змішування різних за походженням культур — R1b та кавказької G2a. Саме G2a-L13 є супутником R1b у Європі, У Центральній та Південній Азії. На Балканах, у Дунайському водосточищі та у Центральній Європі частота G2a-L13 дещо пропорційна до відсотка R1b.
Бронзові речі вперше з'явилися у майкопській культурі й згодом поширилися з Передкавказзя у степи до ямної.
Найстародавніший (3300 років до Р. Х.) меч, що був знайдений у 31-й могилі урочища Клади у станиці Новосвободної в Адигеї належить до майкопської культури. Цей меч є менш витонченим, проте схожим на довгі кельтські мечі, що були широко поширені у Європі.
Майкопська культура принесла у східноєвропейський степ з Месопотамії критий віз. Згодом, приблизно у 2000 році до Р. Х. з'являються військові колісниці. Це призвело до успішного, швидкого, переможного просування степової спільноти R1b просторами Євразії. У цьому зв'язку R1b можуть претендувати на роль міфічних «народів моря», що тероризували узбережжя Східного Середземномор'я у 2 тисячоріччі до Р.Х…
Доба ямної культури
За часу ямної культури (3500-2500 роки до Р. Х.) R-M269 жили та мішалися з першоприбулими R1a. За даними лінгвістів саме у цей період формується як лінгва-франка прото-індоєвропейська мова. Подібності прото-індоєвропейської мови з кавказькими та хуритською мовами Західної Азії з одної сторони та уральських мов Волго-Уральського краю з іншої вказують на понто-каспійський степ як на важливий регіон її походження. Також за часу ямної культури поширилися криті вози, як засіб пересувного життя, що згодом допомогло їм заселити широкі простори Євразії. Цими возами продовжували користуватися за доби Римської імперії германські племена та у Галлії.
Підгалузь M269 гаплогрупа R-L23 (R-Z2103) була знайдена превалентною у стародавніх зразках що відносяться до ямної культури.
Західна частина простору ямної культури від Дону до Дністра й згодом до Подунав'я була найгустіше населена племенами R1b з меншістю у 5-10 % R1a. Згодом ця західна спільнота мігрувала на Балкани й уздовж Дунаю у Західну Європу, а також на південь у Грецію та Анатолію — хвилі дорійців, хеттів, фригійців та мушків (пращурів вірмен).
Східна частина ямної культури у Надволжі та Надураллі навпаки була переважне населена R1a з відчутною меншістю R1b. Східна спільнота ямної культури разом з R1a поширилася на схід у Центральну Азію до Алтаю та Південного Сибіру, Східного Туркестану та Китаю; на Південь до Афганістану, Пакистану, Індії та Туркменії. Вони залишили нащадків серед меншості башкир (М269 та М478), уйгур, туркмен, казахів, чувашів, балкарців, алтайців та інших народів Центральної та Південної Азії..
Північні частини ямної культури у лісостепу України та Росії були населені R1a з малочисельними R1b. Так у носіях культури шнурової кераміки а також сучасних українців, білорусів та росіян R1a у 7-9 раз чисельніші за носіїв R1b. Північна частина ямної культури з лісостепу просунулася на північ, доєднуючися до культури шнурової кераміки R1a поширилися у східній Європі та навколо Балтики у Польщі, Прибалтиці, Скандинавії та північній Німеччині.
R-Z2103 та хетти
Хетти постали через декілька сторіч після зникнення майкопської та ямної культур. Частина племен ямників Z2103 обминули Чорне море з заходу залишив невеликий слід у східній Європі, та помітний — на Балканах та оселилися в Анатолії. Z2103 сьогодні є найпоширенішою формою індоєвропейських галузей R1b у Туреччині.
Ямна культура у понтійсько-каспійському степу змінилася зрубною культурою (1600—1200 роки до Р. Х.), що відобразила рух з східноєвропейського лісостепу R1a-Z282 на південь.
Половецько-козацька R-М478 у Центральній Азії та Україні
R-M478 є «азійською», "кипчацькою" й у випадку України - "половецько-козацькою" галуззю гаплогрупи R-P297.
M73 та М478 обминули Каспій з Півночі й заселили Туран. За іншою версією потрапили у Туран через сучасну території Азербайджану, Ірану та Туркменістану. Згодом були відомі як саки. Носії R-M478 були носіями давніх індоєвропейських мов Центральної Азії й тюркською експансією стали носіями переважно тюркських мов. З тюркізацією Центральної Азії стали відомі як кипчаки, половці, кумани. В Україні представники гаплогрупи належать до нащадків запорозьких козаків.
Археологічні зразки гаплогрупи R1b
- людина з Віллабруна-1 (або 1,215,433), що належить епіграветській культурі з Кісмонської долини у італійській провінції Венето, що жив 14 тисяч років тому, й належав до R1b1a* (R-L754*)[33];
- чоловік-«самарський мисливець-збирач» (I0124; SVP44; M340431), що жив приблизно 7 тисяч років тому над Середньою Волгою зі зразком R1b1* (R-L278*).[34]
Таблиця
| Регіон / Країна | Зразки | R-M269 | R-U106 | R-U106-1 |
|---|---|---|---|---|
| Австрія[35] | 22 | 27 % | 23 % | 0.0 % |
| Латинська Америка[35] | 33 | 0.0 % | 0.0 % | 0.0 % |
| Чехія[35] | 36 | 28 % | 14 % | 0.0 % |
| Данія[35] | 113 | 34 % | 17 % | 0.9 % |
| Східна Європа[35] | 44 | 5 % | 0.0 % | 0.0 % |
| Англія[35] | 138 | 57 % | 20 % | 1.4 % |
| Франція[35] | 56 | 52 % | 7 % | 0.0 % |
| Німеччина[35] | 332 | 43 % | 19 % | 1.8 % |
| Ірландія[35] | 102 | 80 % | 6 % | 0.0 % |
| Італія[36] | 34 | 53 % | 6 % | 0.0 % |
| Йорданія[35] | 76 | 0.0 % | 0.0 % | 0.0 % |
| Близький Схід[35] | 43 | 0.0 % | 0.0 % | 0.0 % |
| Нідерланди[35] | 94 | 54 % | 35 % | 2.1 % |
| Океанія[35] | 43 | 0.0 % | 0.0 % | 0.0 % |
| Оман[35] | 29 | 0.0 % | 0.0 % | 0.0 % |
| Пакистан[35] | 177 | 3 % | 0.0 % | 0.0 % |
| Палестина[35] | 47 | 0.0 % | 0.0 % | 0.0 % |
| Польща[35] | 110 | 23 % | 8 % | 0.0 % |
| Росія[35] | 56 | 21 % | 5.4 % | 1.8 % |
| Словенія[35] | 105 | 17 % | 4 % | 0.0 % |
| Швейцарія[35] | 90 | 58 % | 13 % | 0.0 % |
| Туреччина[35] | 523 | 14 % | 0.4 % | 0.0 % |
| Україна[35] | 32 | 25 % | 9 % | 0.0 % |
| США[35] | 58 | 5 % | 5 % | 0.0 % |
| США (білі) | 125 | 46 % | 15 % | 0.8 % |
| США (негри) | 118 | 14 % | 2.5 % | 0.8 % |
Примітки
- International Society of Genetic Genealogy (ISOGG) — Y-DNA Haplogroup R and its Subclades
- Ornella Semino, A. Silvana Santachiara-Benerecetti, Francesco Falaschi, L. Luca Cavalli-Sforza and Peter A. Underhill, "Ethiopians and Khoisan Share the Deepest Clades of the Human Y-Chromosome Phylogeny, " The American Journal of Human Genetics, Volume 70, Issue 1, 265—268, 1 January 2002.
- R. Gonçalves, A. Freitas, M. Branco, A. Rosa, A. T. Fernandes, L. A. Zhivotovsky, P. A. Underhill, T. Kivisild, A. Brehm: Y-chromosome lineages from Portugal, Madeira and Açores record elements of Sephardim and Berber ancestry. In: Annals of human genetics. Band 69, Pt 4Juli 2005, S. 443–454, ISSN 0003-4800. DOI:10.1111/j.1529-8817.2005.00161.x. PMID 15996172.
- V. Grugni, V. Battaglia, B. Hooshiar Kashani, S. Parolo, N. Al-Zahery, A. Achilli, A. Olivieri, F. Gandini, M. Houshmand, M. H. Sanati, A. Torroni, O. Semino: Ancient migratory events in the Middle East: new clues from the Y-chromosome variation of modern Iranians. In: PloS one. Band 7, Nummer 7, 2012, S. e41252, ISSN 1932-6203. DOI:10.1371/journal.pone.0041252. PMID 22815981. Повний текст на PMC: 3399854.
- Балаганская О. А. Полиморфизм Y-хромосомы у тюркоязычного населения Алтая, Саян, Тянь-Шаня и Памира в контексте взаимодействия генофондов Западной и Восточной Евразии // Автореферат кандидатской диссертации по биологическим наукам. М., МГНЦ РАМН, 2011, С.10.
- C. Capelli, F. Brisighelli, F. Scarnicci, B. Arredi, A. Caglia', G. Vetrugno, S. Tofanelli, V. Onofri, A. Tagliabracci, G. Paoli, V. L. Pascali: Y chromosome genetic variation in the Italian peninsula is clinal and supports an admixture model for the Mesolithic-Neolithic encounter. In: Molecular phylogenetics and evolution. Band 44, Nummer 1, Juli 2007, S. 228–239, DOI:10.1016/j.ympev.2006.11.030, PMID 17275346.
- Estimating Scandinavian and Gaelic Ancestry in the Male Settlers of Iceland — Agnar Helgason et al., 2000, Am. J. Hum. Genet. 67:697-717, 2000
- https://www.familytreedna.com/groups/ukrainian-dna/dna-results
- https://www.familytreedna.com/groups/ukraine/dna-results
- https://www.familytreedna.com/groups/russiadna/dna-results
- Massive migration from the steppe is a source for Indo-European languages in Europe, Haak et al, 2015
- Population genomics of Bronze Age Eurasia, Allentoft et al, 2015
- Eight thousand years of natural selection in Europe, Mathieson et al, 2015
- Myres, Natalie M.; Rootsi, Siiri; Lin, Alice A.; Järve, Mari; King, Roy J.; Kutuev, Ildus; Cabrera, Vicente M.; Khusnutdinova, Elza K. та ін. (January 2011). A major Y-chromosome haplogroup R1b Holocene era founder effect in Central and Western Europe. European Journal of Human Genetics (англ.) 19 (1). с. 95–101. ISSN 1018-4813. doi:10.1038/ejhg.2010.146. Процитовано 21 жовтня 2017.
- Karafet, Tatiana M.; Mendez, Fernando L.; Meilerman, Monica B.; Underhill, Peter A.; Zegura, Stephen L.; Hammer, Michael F. (2008-5). New binary polymorphisms reshape and increase resolution of the human Y chromosomal haplogroup tree. Genome Research 18 (5). с. 830–838. ISSN 1088-9051. PMC PMC2336805. PMID 18385274. doi:10.1101/gr.7172008. Процитовано 21 жовтня 2017.
- Semino, O.; Passarino, G; Oefner, PJ; Lin, AA; Arbuzova, S; Beckman, LE; De Benedictis, G; Francalacci, P; Kouvatsi, A та ін. (2000). The genetic legacy of paleolithic Homo sapiens sapiens in extant Europeans: a Y chromosome perspective. Science 290 (5494): 1155–59. PMID 11073453. doi:10.1126/science.290.5494.1155.
- Balaresque, Patricia; Bowden, Georgina R.; Adams, Susan M.; Leung, Ho-Yee; King, Turi E. та ін. (2010). A Predominantly Neolithic Origin for European Paternal Lineages. У Penny, David. PLOS Biology (Public Library of Science) 8 (1): e1000285. PMC 2799514. PMID 20087410. doi:10.1371/journal.pbio.1000285. Процитовано 19 серпня 2014.
- Sjödin, Per; François, Olivier (2011). Wave-of-Advance Models of the Diffusion of the Y Chromosome Haplogroup R1b1b2 in Europe. У Lalueza-Fox, Carles. PLoS ONE 6 (6:e21592): e21592. PMC 3123369. PMID 21720564. doi:10.1371/journal.pone.0021592.
- B. Arredi; E. S. Poloni; C. Tyler-Smith (2007). The peopling of Europe. У Crawford, Michael H. Anthropological genetics: theory, methods and applications. Cambridge, UK: Cambridge University Press. с. 394. ISBN 0-521-54697-4.
- Morelli, Laura; Contu, Daniela; Santoni, Federico; Whalen, Michael B.; Francalacci, Paolo; Cucca, Francesco (2010). A Comparison of Y-Chromosome Variation in Sardinia and Anatolia Is More Consistent with Cultural Rather than Demic Diffusion of Agriculture. У Lalueza-Fox, Carles. PLoS ONE 5 (4): e10419. PMC 2861676. PMID 20454687. doi:10.1371/journal.pone.0010419.
- http://www.pnas.org/content/95/15/9053.full.pdf+html
- Maglio, Michael R. (14 серпня 2014). Biogeographical Evidence for the Iberian Origins of R1b-L278 via Haplotype Aggregation (2014). Origins DNA. Процитовано 12 листопада 2014.
- Chiaroni, J; Underhill, P; Cavalli-Sforza, L.L. (2009). Y chromosome diversity, human expansion, drift and cultural evolution. PNAS 106 (48): 20174:20179. PMC 2787129. PMID 19920170. doi:10.1073/pnas.0910803106.
- Cruciani et al.; Trombetta, Beniamino; Antonelli, Cheyenne; Pascone, Roberto; Valesini, Guido; Scalzi, Valentina; Vona, Giuseppe; Melegh, Bela та ін. (2010). Strong intra- and inter-continental differentiation revealed by Y chromosome SNPs M269, U106 and U152. Forensic Science International: Genetics 5 (3): e49. PMID 20732840. doi:10.1016/j.fsigen.2010.07.006.
- Grugni, Viola; Battaglia, Vincenza; Kashani, Baharak Hooshiar; Parolo, Silvia; Al-Zahery, Nadia; Achilli, Alessandro; Olivieri, Anna; Gandini, Francesca та ін. (18 липня 2012). Ancient Migratory Events in the Middle East: New Clues from the Y-Chromosome Variation of Modern Iranians. PLOS ONE 7 (7). с. e41252. ISSN 1932-6203. doi:10.1371/journal.pone.0041252. Процитовано 20 жовтня 2017.
- Category: Y-DNA Haplogroup R-M335. www.wikitree.com (англ.). Процитовано 21 жовтня 2017.
- Кластеры внутри R1b1b1-M73. forum.molgen.org. Процитовано 20 жовтня 2017.
- Разбор случая kodak. forum.molgen.org. Процитовано 20 жовтня 2017.
- R-M478 YTree. www.yfull.com. Процитовано 7 травня 2020.
- ISOGG, Y-DNA Haplogroup R and its Subclades — 2017 (1 August 2017).
- https://www.eupedia.com/europe/Haplogroup_R1b_Y-DNA.shtml
- https://www.biorxiv.org/content/early/2017/05/09/135616
- Fu, Qiaomei et al. «The genetic history of Ice Age Europe». Nature 534, 200—205 (09 June 2016) doi:10.1038/nature17993
- Haak, W.; Lazaridis, I.; Patterson, N.; Rohland, N.; Mallick, S.; Llamas, B.; Brandt, G.; Nordenfelt, S.; Harney, E.; Stewardson, K.; Fu, Q.; Mittnik, A.; Bánffy, E.; Economou, C.; Francken, M.; Friederich, S.; Pena, R. G.; Hallgren, F.; Khartanovich, V.; Khokhlov, A.; Kunst, M.; Kuznetsov, P.; Meller, H.; Mochalov, O.; Moiseyev, V.; Nicklisch, N.; Pichler, S. L.; Risch, R.; Rojo Guerra, M. A. та ін. (2015). Massive migration from the steppe was a source for Indo-European languages in Europe. Nature 522: 207–211. Bibcode:2015Natur.522..207H. PMC 5048219. PMID 25731166. doi:10.1038/nature14317.
- Myres, NM; Ekins, JE; Lin, AA; Cavalli-Sforza, LL; Woodward, SR; Underhill, PA (2007). Y-chromosome Short Tandem Repeat DYS458.2 Non-consensus Alleles Occur Independently in Both Binary Haplogroups J1-M267 and R1b3-M405. Croatian medical journal 48 (4): 450–9. PMC 2080563. PMID 17696299.
- Myres, Natalie; Rootsi, Siiri; Lin, Alice A; Järve, Mari; King, Roy J; Kutuev, Ildus; Cabrera, Vicente M; Khusnutdinova, Elza K та ін. (2010). A major Y-chromosome haplogroup R1b Holocene effect in Central and Western Europe. European Journal of Human Genetics 19 (1): 95–101. PMC 3039512. PMID 20736979. doi:10.1038/ejhg.2010.146.
Відомі носії
- Микола ІІ (Романов) (1868—1918) — російський імператор (1894—1917)
- Микола Коперник (1473—1543) — польський астроном та математик
- Вудро Вільсон (1856—1924) — 28-ий американський президент,
- Чарльз Дарвін (1809—1882) — еволюціонист
- Тутанхамон (14 сторіччя до Р. Х.) — єгипетський фараон
 Романов
Романов Коперник
Коперник Вилсон
Вилсон Дарвин
Дарвин Тутанхамон
Тутанхамон
Джерела
- Balaresque, Patricia; Bowden, Georgina R.; Adams, Susan M.; Leung, Ho-Yee; King, Turi E.; et al. (2010). Penny, David, ed. «A Predominantly Neolithic Origin for European Paternal Lineages». PLOS Biology. Public Library of Science. 8 (1): e1000285. PMC 2799514 Freely accessible. PMID 20087410.
- Zalloua, PA; Xue, Y; Khalife, J; Makhoul, N; Debiane, L; Platt, DE; Royyuru, AK; Herrera, RJ; Hernanz, DF; et al. (2008). «Y-Chromosomal Diversity in Lebanon Is Structured by Recent Historical Events». American Journal of Human Genetics. 82 (4): 873–82. PMC 2427286 Freely accessible. PMID 18374297.
- Myres, NM; Ekins, JE; Lin, AA; Cavalli-Sforza, LL; Woodward, SR; Underhill, PA (2007). «Y-chromosome Short Tandem Repeat DYS458.2 Non-consensus Alleles Occur Independently in Both Binary Haplogroups J1-M267 and R1b3-M405». Croatian medical journal. 48 (4): 450–9. PMC 2080563 Freely accessible. PMID 17696299.