Келихоподібні клітини
Келихоподібні[2][3], або бокалоподі́бні[4] кліти́ни — епітеліальні залозисті клітини слизових оболонок тварин, які виділяють білковий компонент слизу. У людини і хребетних тварин поодинокі або численні келихоподібні клітини розташовані в поверхневому шарі багатьох слизових оболонок (травний тракт, дихальні шляхи, волога оболонка ока), у риб, амфібій та деяких безхребетних — і в епідермісі шкіри.
| Бокалоподібна клітина | |
|---|---|
 | |
| Деталі | |
| Ідентифікатори | |
| Латина | exocrimohsinocytus caliciformis |
| MeSH | D020397 |
| TH | H3.04.03.0.00009, H3.04.03.0.00016 і H3.05.00.0.00006 |
| FMA | 13148[1] |
| Анатомічна термінологія | |
Будова
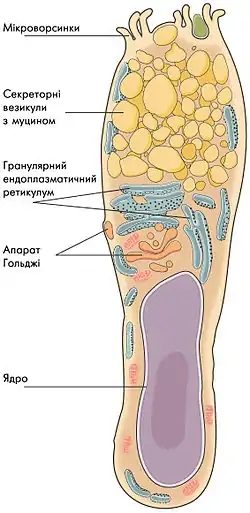
Келихоподібні клітини знаходяться в одношаровому епітелії слизових оболонок. Клітини мають видовжену форму із розширенням до верхівки, чим нагадують келих чи бокал. Верхня (апікальна) частина спрямована до поверхні епітелію і містить численні секреторні гранули, наповнені муцином, які підвищують схожість з келихом. У тоншій базальній частині знаходиться ядро, а вище нього — апарат Гольджі.
Секреторна функція
Основним елементом слизу, який секретують келихоподібні клітини, є муцини — глікопротеїни з високою молекулярною масою. Гени муцинів експресуються в ядрі бокалоподібної клітини постійно, а також регулюються багатьма факторами, зокрема прозапальними інтерлейкінами.[джерело?] Синтезований пептид муцину в ендоплазматичному ретикулумі підлягає N-глікозилюванню, а звідти спрямовується везикулами до апарату Гольджі, де модифікується шляхом O-глікозилювання N-ацетилгалактозаміном серинового та треонінового залишків центральних доменів білка. В апараті Гольджі додатково до N-ацетилгалактозаміну приєднуються гілочки інших моносахаридів. Зрілий білок містить 80% вуглеводного компоненту за сухою масою. Він надходить у секреторних гранулах до верхньої частини бокалоподібної клітини.[5]
Секреторна гранула являє собою мембранну везикулу, всередині якої багато іонів кальцію та кислий pH близько 6, що дозволяє дуже щільно упакувати молекули муцину. Окремі молекули муцину тримеризуються своїми амінокінцями за участі іону кальцію, а вільні C-кінці білка з'єднуються між собою, утворюючи сітку з шестигранними комірками. Надлишок іонів водню призводить до стиснення цієї сітки шляхом складання паралельними шарами. Після вивільнення везикули відбувається розширення її вмісту в 1000 разів.[6]
Секреція муцину можлива двома шляхами: або шляхом екзоцитозу, коли везикули зливаються з мембраною та вивільняють свій вміст у позаклітинний простір, або шляхом злиття усіх везикул в одну та вивільнення їх назовні шляхом руйнації келихоподібної клітини.[6] При ініціації екзоцитозу активується Ca2+/K+ обмінник мембрани везикули, та іони кальцію починають викачуватися у цитоплазму.[7]
Регуляція секреції
Секреція муцину залежить від багатьох факторів. Для вивільнення муцину в кишечнику обов'язково мають працювати компоненти системи автофагії та генеруватися ендогенні вільні радикали.[8] Ацетилхолін збільшує секрецію бокалоподібних клітин у кишечнику різних видів ссавців, гістамін стимулює їх секрецію в товстому кишечнику, але не в тонкому. Різний вплив на секрецію муцинів здійснюють простагландини.[6]
Інші функції
Бокалоподібні клітини кишечника здатні до спонтанного захоплення антигенів та до антиген-презентації.[9]
Розвиток
У кишечнику бокалоподібні клітини походять разом з ентерохромафінними клітинами з LGR5-позитивних стовбурових клітин. Ці стовбурові клітини дають початок проміжним стовбуровим клітинам, що швидко діляться і є попередниками диференційованих епітеліальних клітин кишечника, зокрема й бокалоподібних. Сформована бокалоподібна клітина живе 3-7 днів.[10]
Поширення
Бокалоподібні клітини ссавців поширені на багатьох слизових оболонках. Вони знаходяться в одношаровому епітелії серед інших епітеліальних клітин. Вони наявні впродовж усього травного тракту: в ротовій порожнині, глотці, стравоході, шлунку, тонкому й товстому кишечнику. Товстий кишечник ссавців має значно більше бокалоподібних клітин аніж тонкий.[6] У шлунку секрецію слизу забезпечують в основному додаткові клітини, які відрізняються від бокалоподібних морфологічно. Також бокалоподібні клітини функціонують у повітроносних шляхах: в епітелії носової порожнини, в гортані, трахеї, бронхах і бронхіолах. У безхрящових бронхіолах вони є єдиними виробниками слизу.[7] Келихоподібні клітини знаходяться в епітеліальному шарі кон'юнктиви ока, де беруть участь у формуванні зволожувального секрету.[11]
Особливості
Тонкий кишечник
У тонкому кишечнику бокалоподібні клітини секретують муцин MUC2, який спочатку кріпиться до апікальної мембрани клітини. Надалі спеціальна металопротеаза ентероцитів мепрін β відщеплює цей муцин від поверхні. Мепрін знаходиться на поверхні ентероцита і відокремлюється від нього при появі бактерій. Тому в тварин, яких тримають у стерильних умовах, слиз у тонкому кишечнику міцно тримається на поверхні клітин.[6] Окрім муцину MUC2 гранули клітин тонкого кишечнику містять білки FCGBP, CLCA1, ZG16, та AGR2. Для виділення муцину бокалоподібні клітини потребують секреції бікарбонату сусідніми ентероцитами.
Товстий кишечник
У товстому кишечнику бокалоподібні клітини мають власний білок-транспортер бікарбонату Бестрофін 2.
Захворювання
Масова загибель келихоподібних клітин трапляється при кератокон'юнктивіті, або хворобі сухих очей. Бокалоподібні клітини гинуть шляхом апоптозу, викликаного цитокінами γ-інтерфероном та інтерлейкінами.[11]
Гіперплазія келихоподібних клітин поширена при астмі та хронічному обструктивному захворюванні легень. Вона виражається у збільшенні кількості клітин, зміні їх форми, посиленні секреції муцинів.[7]
Секреторні клітини риб та амфібій
В епідермісі нижчих хребетних, таких як круглороті, риби та амфібії присутні секреторні клітини, які схожі на келихоподібні клітини формою, мікроструктурою, наявністю великих секреторних везикул. Вважається, що гранули келихоподібних клітин шкіри шпоркових жаб містять муцини, хоча склад їх не досліджений. Відомо, що їхній секрет містить лектин Xeel, а в самих клітинах було визначено глікопротеїн хондроїтин сульфат.[12] У риб келихподібні клітини реагують на присутність бактерій у воді збільшенням секреції та більш активним глікозилюванням білків слизу. Передбачається, що механізми активації секреції під дією бактерій у них можуть бути подібні до таких у ссавців.[13]
Примітки
- Foundational Model of Anatomy
- Ганонг, Вільям Ф. (2002). Фізіологія людини (Переклад з англ.). Львів: БаК. с. 784. ISBN 0-8385-8282-6.
- М. Е. Дзержинський, Г. В. Островська, Н. В. Скрипник, С. М. Гарматіна (2012). Гістологія. Практикум. Київ: Видавничо-поліграфічний центр "Київський університет". с. 88.
- під ред. Широбоков В. П. (2011). Медична мікробіологія, вірусологія та імунологія (вид. 2). Вінниця: Нова Книга. с. 952. ISBN 978-966-382-325-6.
- Thaher Pelaseyed, Joakim H. Bergstrom, Jenny K. Gustafsson, Anna Ermund, George M. H. Birchenough, Andre Schutte, Sjoerd van der Post, Frida Svensson, Ana M. Rodriguez-Pineiro, Elisabeth E. L. Nystrom, Catharina Wising, Malin E. V. Johansson, Gunnar C. Hansson (July 2014). The mucus and mucins of the goblet cells and enterocytes provide the first defense line of the gastrointestinal tract and interact with the immune system. Immunological reviews 260 (1): 8–20. PMID 24942678. doi:10.1111/imr.12182.
- G. M. H. Birchenough, M. Ev Johansson, J. K. Gustafsson, J. H. Bergstrom & G. C. Hansson (July 2015). New developments in goblet cell mucus secretion and function. Mucosal immunology 8 (4): 712–719. PMID 25872481. doi:10.1038/mi.2015.32.
- Duncan F. Rogers (January 2003). The airway goblet cell. The international journal of biochemistry & cell biology 35 (1): 1–6. PMID 12467641.
- Patel, Khushbu K; Miyoshi, Hiroyuki; Beatty, Wandy L; Head, Richard D; Malvin, Nicole P; Cadwell, Ken; Guan, Jun-Lin; Saitoh, Tatsuya; Akira, Shizuo; Seglen, Per O; Dinauer, Mary C; Virgin, Herbert W; Stappenbeck, Thaddeus S (2013). Autophagy proteins control goblet cell function by potentiating reactive oxygen species production. The EMBO Journal 32 (24): 3130–3144. ISSN 02614189. doi:10.1038/emboj.2013.233.
- McDole, Jeremiah R.; Wheeler, Leroy W.; McDonald, Keely G.; Wang, Baomei; Konjufca, Vjollca; Knoop, Kathryn A.; Newberry, Rodney D.; Miller, Mark J. (2012). Goblet cells deliver luminal antigen to CD103+ dendritic cells in the small intestine. Nature 483 (7389): 345–349. ISSN 0028-0836. doi:10.1038/nature10863.
- Barker, Nick (2013). Adult intestinal stem cells: critical drivers of epithelial homeostasis and regeneration. Nature Reviews Molecular Cell Biology 15 (1): 19–33. ISSN 1471-0072. doi:10.1038/nrm3721.
- Zhang, Xiaobo; De Paiva, Cintia S.; Su, Zhitao; Volpe, Eugene A.; Li, De-Quan; Pflugfelder, Stephen C. (2014). Topical interferon-gamma neutralization prevents conjunctival goblet cell loss in experimental murine dry eye. Experimental Eye Research 118: 117–124. ISSN 00144835. doi:10.1016/j.exer.2013.11.011.
- Dubaissi, E.; Rousseau, K.; Lea, R.; Soto, X.; Nardeosingh, S.; Schweickert, A.; Amaya, E.; Thornton, D. J. та ін. (2014). A secretory cell type develops alongside multiciliated cells, ionocytes and goblet cells, and provides a protective, anti-infective function in the frog embryonic mucociliary epidermis. Development 141 (7): 1514–1525. ISSN 0950-1991. doi:10.1242/dev.102426.
- van der Marel, M; Caspari, N; Neuhaus, H; Meyer, W; Enss, M-L; Steinhagen, D (2010). Changes in skin mucus of common carp,Cyprinus carpioL., after exposure to water with a high bacterial load. Journal of Fish Diseases 33 (5): 431–439. ISSN 01407775. doi:10.1111/j.1365-2761.2010.01140.x.
Джерела
- R. D. Specian & M. G. Oliver (February 1991). Functional biology of intestinal goblet cells. The American journal of physiology 260 (2 Pt 1): C183–C193. PMID 1996606.
- Duncan F. Rogers (January 2003). The airway goblet cell. The international journal of biochemistry & cell biology 35 (1): 1–6. PMID 12467641.