Звід (органела)
Звід (англ. vault) — еукаріотична органела, що хімічно являє собою рибонуклеопротеїн. Під електронним мікроскопом ці органели нагадують звід (склепіння) купола собору з віссю симетрії 39-го порядку[1]. Функції зводу є погано вивченими, проте на сьогодні є відомості про їх участь у різних сигнальних шляхах клітини. Можливо, зводи залучені до розвитку явища множинної медикаментозної стійкості до протипухлинної хіміотерапії. Вони присутні у багатьох типах еукаріотичних клітин і є висококонсервативними серед еукаріотів[2].

Історія вивчення
Звід було відкрито і успішно виділено з печінки миші у 1986 році клітинним біологом Ненсі Кедерша (англ. Nancy Kedersha) і біохіміком Леонардом Ромом (англ. Leonard Rome) з медичної школи Каліфорнійського університету у Лос-Анджелесі[3]. Спочатку зводи були описані як яйцеподібні часточки, що забруднювали препарати окаймлених клатрином везикул. Часточки були виділені за допомогою центрифугування у градієнті щільності сахарози і електрофорезу у агарозному гелі. Виявилось, що вони мають симетричну бочкоподібну структуру, схожу на склепіння готичного собору, за що часточки і отримали свою назву (від англ. vault — звід). Спочатку розмір зводу оцінили у 35 × 35 × 65 нм³, але згодом з використанням кріоелектронної мікроскопії він був уточнений до 41 × 41 × 72,5 нм³. Таким чином, зводи є найбільшими з коли-небудь описаних цитозольних неікосаедричних нуклепротеїнів. Надалі структура зводів вивчалась за допомогою рентгеноструктурного аналізу і ядерного магнітного резонансу. У 2009 році структуру зводів з печінки миші було визначено з розширенням 3,5 Å[4].
Структура зводу
Зводи — це найбільші рибонуклеопроїнові частинки. За розміром вони перевершують рибосому і важать близько 13 МДа[5]. Зводи майже повністю складаються з білків, через що їх складно забарвлювати стандартними методами. Білкова частина зводу представлена багатьма молекулами головного білка зводу(MVP) (95,8 кДа), на частку якого припадає більше 70 % усіх білків у частинці[6], а також білками VPARP (∼192 кДа) s TEP1 (∼291 кДа). Окрім цього, до складу зводу входять зводові РНК (зРНК) довжиною 86—141 нуклеотидів[7]. Сумарну масу РНК у зводі оцінюють у ∼460 кДа[4].
Часточка зводу досягає близько 670 Å у довжину і має найбільшій діаметр ∼400 Å. Стінка має товщину всього 15–25 Å; всередині неї знаходиться порожнина довжиною близько 620 Å і максимальним діаметром ∼350 Å. Частинка складається з двох симетричних половин, кожна з яких складається з трьох частин: тіла, плечової ділянки і шапочки. Тіло містить 78 копій 9 повторюваних структурних доменів MVP (39 копій у кожній половині), звужена частина сформована зв'язаними кінець до кінця структурними повторюваними доменами R1. Висота плечової ділянки складає ∼25 Å, а діаметр ∼315 Å. Шапочки присутні обабіч частинки, і кожна з них містить 39 копій домену шапочка-спіраль (амінокислотні залишки MVP з Асп647 по Лей802) і домену шапочка-коло (з Глі803 по Ала845). Висота шапочки складає ∼155 Å, а внутрішній і зовнішній діаметри домену шапочка-коло досягають ∼50 Å і ∼130 Å відповідно[4].
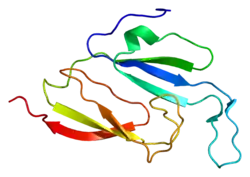
MVP складається з 9 повторюваних структурних доменів (R1—R9). Домени R8 і R9 складаються з п'яти антипаралельних β-листів, що позначаються S1, S2, S3, S4 і S5. У решти семи доменах є два додаткових β-листи (S2a і S2b), вставлених між S2 і S3. За деякими даними, R1, як і R8 і R9, складається з п'яти антипаралельних β-листів, у той час як у R2 є два довших антипаралельних листки між S2 і S3. Кожний домен має гідрофобний ефект. Аналіз амінокислотних послідовностей показав, що у R3 і R4, можливо, є два домени типу «EF hand». Подальше вивчення показало, що MVP взаємодіє з іншими білками, такими як PTEN, через ймовірні два домени EF hand за участю іонів Кальцію, втім, з цим узгоджуються не усі експериментальні дані[4].
Плечова ділянка (з Pro520 по Val646) згортається у один α/β-глобулярний домен з 4 антипаралельними бета-листами з одного боку і з 4 α-спіралями з іншого. Очевидно, що саме у плечовій ділянці знаходяться елементи, що відповідають за взаємодію зводу з ліпідними рафтами[4].
Домен шапочка-спіраль згортається в α-спіраль із 42 витків, яка укладається у суперспіраль. Домен шапочка-коло розташований на кінці шапочки і формує U-подібну структуру зі спіральними елементами на обох кінцях[4].
зРНК розташовуються у шапочках на кінцях часточок зводу. Білок TEP1, очевидно, розташовується згори плоскої частини шапочки, де його ділянка WD40 repeat формує структуру типу кільцевого β-пропелера. N-кінцева частина TEP1 містить 4 повторюваних домени незрозумілого функціонального призначення, РНК-зв'язувальний домен і АТФ/ГТФ-зв'язувальний домен. Було показано, що TEP1 взаємодіє з теломеразною РНК і різними людськими зРНК. VPARP розташовані здебільшого у шапочці зводу[4].
| Компонент | Характеристика | Локус | Положення і взаємодія у зводі | Функції |
|---|---|---|---|---|
| MVP | 96 копій білку становлять основну масу зводу. Здатні самоскладатись у звід. Не мають функцій поза межами зводу | 16p11.2 | Тіло і шапочка зводу, немає необхідних партнерів | Структурна; керує зв'язуванням із сигнальними білками |
| VPARP | Слабка полі(АДФ-рибоза)-полімераза, лише частково зв'язана зі зводом | 13q11 | Шапочка, безпосередньо зв'язуються з N-кінцевою частиною MVP | Каталітична: полі (АДФ)-рибозилювання MVP |
| TEP1 | Зв'язується з теломеразою і зводом; не необхідний для теломерази | 14q11.2 | Шапочка, безпосереднє зв'язування з MVP | Каталітична: збирання зРНК-мішеней у зводі |
| зРНК | hgv1—3, hgv4 не експресується; лише частково зв'язані зі зводом | 5q33.11
Xp11.22 |
Шапочка, зв'язуються з TEP1 | Неструктурна, функції погано вивчені |
Функції
Широке розповсюдження зводів і їх еволюційна консервативність вказують на вигоду присутності в цих органел важливих біологічних функцій, хоча про них відомо дуже мало. Невідомо нічого про первісні функції зводів у клітинах протистів. Тим не менш, є кілька теорій щодо ролі зводів у клітинах ссавців. Серед них вказується, що особливо велика кількість зводів присутня у клітинах і тканинах, пов'язаних з очищенням організму, наприклад, у макрофагах.
Також було висловлено гіпотезу, що зводи слугують основними «корками» у ядерних порових комплексах. Імунофлуоресцентний аналіз із використанням антитіл проти зводів показав, що в ізольованих ядрах клітин печінки миші зводи розташовуються на поверхні ядерної мембрани. Імуноелектронна мікроскопія, з використанням вторинних антитіл, кон'югованих із Ауромом, показало, що в ізольованих ядрах зводи є асоційованими з ядерними поровими комплексами. Тому, можливо, зводи можуть брати участь у ядерноцитоплазматичному транспорті.
2005 року було висунуто припущення, що людські зРНК hvg1 та hvg2 можуть зв'язуватись із протираковим препаратом мітоксантроном, а також відігравати важливу роль в експорті токсичних сполук. Втім, інше дослідження показало, що порушення роботи гену MVP у мишей не приводило до посилення чутливості до цитостатичних препаратів. Більше того, миші дикого типу та дефектні по MVP миші демонстрували однакову відповідь на доксорубіцин. Під час ще одного дослідження було продемонстровано, що нокдаун MVP за допомогою малих інтерферувальних РНК не впливав на видалення доксорубіцину з ядра. Окрім того, підвищення експресії MVP у хемочутливих клітинах не посилювало стійкості до препаратів. Ці результати свідчать на користь того, що MVP і зводи не вносять безпосереднього вкладу до стійкості до цитостатичних агентів.
Низка досліджень показала участь зводів у різних сигнальних шляхах клітини, і кількість таких шляхів постійно зростає. На дріжджевій двогібридній системі було показано, що MVP може зв'язуватись з PTEN — білком-супресором пухлин, який фосфорилює фосфатидилінозитол-3,4,5-трифосфат, негативно регулюючи сигнальний шлях фосфоінозитид-3-кінази SHP-2, що містить домен SH2 (Src homology 2), і слугує скеффолд-білком у сигнальному шляху епідермального фактору росту (EGF). Виявилось, що SHP-2 зв'язувались з MVP, фосфорильованим по залишках тирозину, і це зв'язування посилилось під дією EGF. Таким чином, MVP функціонує як скеффолд-білок для SHP-2 і кіназ, регульованих внутрішньоклітинно, і регуляція фосфорилювання MVP через SHP-2 може мати важливе значення для виживання клітини. Окрім того, було показано взаємодію між MVP і доменом SH2 Src у клітинах шлунка людини і клітинах раку шлунка 253J. Імунопреципітація і імунофлуорисцентний аналіз показали, що EGF посилював взаємодію між MVP і Src, і вона блокувалась під дією інгібітору Src PP2, EGF також стимулює переміщення MVP з ядра у гіалоплазму і навколоядерну зону цитоплазми, де MVP колокалізується з Src. Є припущення щодо ролі MVP як нового регулятора Src-опосередкованих сигнальних каскадів. Було встановлено, що MVP є білком, індукованим інтерфероном γ (IFN-γ): у відповідь на IFN-γ спостерігалось значне підвищення рівня мРНК і самого білка MVP. Ця активація залучена до взаємодії STAT1 і сайту, що активується IFN-γ, у проксимальному перегляді MVP. Окрім того, IFN-γ значно посилював темпи трансляції MVP. Показано, що зводи можуть взаємодіяти з естрогеновими рецепторами при зв'язуванні з естрадіолом і разом з рецепторами переносяться у ядро. Відповідно до одних з даних, зводи та MVP можуть взаємодіяти з інсуліноподібним фактором росту 1, HIF1A, а також заторкувати два головних процеси репарації дволанцюгових розривів у ДНК: негомологічне з'єднання кінців і гомологічну рекомбінацію. Таким чином, частинки зводів функціонують як центральні платформи взаємодії у клітинних сигнальних каскадах.
VRARP, інший білок, що входить до складу зводів, є полі(АДФ-рибоза)-полімеразою.
Незвичайна структура і своєрідна динаміка зводів, а також їх великі розміри підводять до думки, що, можливо, зводи функціонують як природних наноконтейнерів для ксенобіотиків, нуклеїнових кислот і білків. Проводиться робота з розробки рекомбінантних зводів, а саме із забезпечення взаємодії зводів із поверхневими клітинними рецепторами і внесення до них різноманітних вантажів.
У таблиці узагальнено основні відомості про білки, з якими взаємодіють зводи:
| Білок | Характеристика | Локус | Взаємодія зі зводом | Функції, пов'язані зі зводом |
|---|---|---|---|---|
| PTEN | Білок-супресор пухлин, головна інгібіторна фосфатаза шляху PI3K. Головний субстрат PIP3 | 10q23.3 | MVP (N-кінець), Ca2+-залежна взаємодія | Посилення ядернах функцій PTEN |
| SHP2 | Протеїнтирозинфосфатаза; активатор росту, опосередкованого рецепторними тирозинкіназами (RTK) | 3q13.13 | MVP (залежна від фосфорилювання MVP взаємодія) | Стимулювання EGRF-опосередкованої MAPK-активності |
| Erk2 | Мітоген-тирозинкиназа, яка активізується; головний передавач RTK-опосередкованих сигналів проліферації | 22q11.22 | MVP (залежна від фосфорилювання MVP взаємодія) | Стимулювання EGRF-опосередкованої MAPK-активності |
| SRC | (Прото)онкоген, тирозинкіназа | 20q11.2 | MVP (залежна від фосфорилювання MVP взаємодія) | Стимулювання EGRF-опосередкованої MAPK-активності |
| COP1 | E3 убіквітинлігаза, у хребетних знищує c-Jun і p53 | 1q25.1—1q25.2 | MVP (фосфорилювання MVP, що зменшується від УФ-випромінення) | Пригнічує EGF-опосередковану активацію MAPK |
| Естрогеновий рецептор | Ядерний рецептор гормону естрадіолу, індукує ліган-залежну транскрипцію генів-мішеней | 6q25.1 | MVP (гормонзалежна взаємодія) | Ядерний імпорт і активація естрогенового рецептора |
| La РНК-зв'язуючий білок | Зв'язує і захищає 3'-кінцеві елементи UUU(OH) транскриптів, синтезованих РНК-полімеразою III | 2q31.1 | зРНК | Захист зРНК |
Клінічне значення
Рак
У 1990-х роках появились повідомлення про те, що зводи можуть безпосередньо бути залучені до розвитку множинної медикаментозної стійкості у ракових клітинах. Виявилось, що білок, пов'язаний з множинною стійкістю і відомий під назвою LRP (англ. Lung Resistance-related Protein — ,білок, пов'язаний з множинною стійкістю у легенях) насправді є людським MVP. Під час проведення другого дослідження зв'язок між зводами і множинною медикаментозною стійкістю був показаний на клітинах людського раку товстої кишки SW-620. Обробка SW-620 бутиратом натрію збільшувала експресію MVP і приводила до стійкості до доксорубіцину, вінкристину, граміцидину D і паклітакселу. Трансфекція клітин рибозимами, специфічними до MVP, пригнічувала ці активності.
Свій вклад у розвиток множинної медикаментозної стійкості можуть вносити і зРНК. 2009 року було встановлено, що некодуючі зРНК можуть процесуватись на малі зРНК (svРНК) за участі «Dicer», які далі функціонують шляхом РНК-інтерференції подібно до мікроРНК: svРНК зв'язуються з білком родини Argonaute і негативно регулюють експресію CYP3A4 — ферменту, що бере участь у метаболізмі ксенобіотиків.
В останні роки накопичуються свідчення того, що зводи пов'язані з функціонуванням систем репарації ДНК у клітині, тому, можливо, вони вносять свій вклад до нечутливості не тільки до хіміотерапії, але й до променевої терапії раку.
Інфекційні захворювання
2007 року дві дослідницькі групи повідомили про участь зводів у відповіді на інфекції. Виявилось, що у людських B-лімфоцитах, інфікованих вірусом Епштейна—Барр, спостерігались підвищені рівні зРНК, які, можливо, якось причетні до захисту від вірусу і/чи до транспортних механізмів. Окрім того, було показано, що при заражені епітеліальних клітин легень людини бактерією Паличкою синьогнійною MVP швидко рекрутувався у ліпідні рафти, де бере участь у механізмах посилення вродженої імунної відповіді. Миші MVP−/− мали у 3,5 рази більше бактерій на грам легеневої тканини, ніж миші дикого типу, і частіше помирали від інфекції, викликаної синьогнійною паличкою.
Еволюційна консервативність
Зводи були описані у ссавців, земноводних, птахів, а також у слизняка Dictyostelium discoideum[2]. Згідно з інформацією із бази даних Pfarm, гомологи білків, що входять до складу зводів, виявлені у інфузорії Paramecium tetraurelia, кінетопластид, багатьох хребетних, актинії Nematostella vectensis, молюсків, пластинчастих, плоских червів (а саме в Ехінокока звичайного) і хоанофлагелятів[8].
У низки еукаріотичних організмів не було виявлено гомологів білків зводів. Серед них такі модельні організми, як рослина Різушка Таля, нематода Caenorhabditis elegans, плодова мушка Дрозофіла чорночерева і пивні дріжджі[9]. Однак, незважаючи на ці винятки, високий ступінь схожості зводів у різних організмів говорить про те, що ці органели мають певне еволюційне значення[2]. Згідно з останніми даними, останній спільний предок еукаріотів мав зводи, проте згодом вони було втрачені у низки груп, серед яких гриби, комахи і, можливо, рослини[10].
Примітки
- PMID 19150846 (PubMed)
Бібліографічний опис з'явиться автоматично через деякий час. Ви можете підставити цитату власноруч або використовуючи бота. - Kedersha, N. L.; Miquel, M. C.; Bittner, D.; Rome, L. H. (1 квітня 1990). Vaults. II. Ribonucleoprotein structures are highly conserved among higher and lower eukaryotes. The Journal of Cell Biology 110 (4). с. 895–901. ISSN 0021-9525. PMC 2116106. PMID 1691193. Процитовано 9 січня 2016.
- PMID 2943744 (PubMed)
Бібліографічний опис з'явиться автоматично через деякий час. Ви можете підставити цитату власноруч або використовуючи бота. - Tanaka H., Tsukihara T. Structural studies of large nucleoprotein particles, vaults.
- Kedersha N. L., Heuser J. E., Chugani D. C., Rome L. H. Vaults.
- PMID 10477748 (PubMed)
Бібліографічний опис з'явиться автоматично через деякий час. Ви можете підставити цитату власноруч або використовуючи бота. - van Zon A., Mossink M. H., Scheper R. J., Sonneveld P., Wiemer E. A. The vault complex.
- Pfam: Family: Vault (PF01505). pfam.xfam.org. Процитовано 9 січня 2016.
- Rome, L.; Kedersha, N.; Chugani, D. (1 серпня 1991). Unlocking vaults: organelles in search of a function. Trends in Cell Biology 1 (2-3). с. 47–50. ISSN 0962-8924. PMID 14731565. Процитовано 9 січня 2016.
- Daly, Toni K.; Sutherland-Smith, Andrew J.; Penny, David (1 січня 2013). In silico resurrection of the major vault protein suggests it is ancestral in modern eukaryotes. Genome Biology and Evolution 5 (8). с. 1567–1583. ISSN 1759-6653. PMC 3762200. PMID 23887922. doi:10.1093/gbe/evt113. Процитовано 9 січня 2016.
Література
- Rome L. H., Kickhoefer V. A. Development of the vault particle as a platform technology. (англ.)
- Reis E. V., Pereira R. V., Gomes M., Jannotti-Passos L. K., Baba E. H., Coelho P. M., Mattos A. C., Couto F. F., Castro-Borges W., Guerra-Sá R. Characterisation of major vault protein during the life cycle of the human parasite Schistosoma mansoni. (англ.)
- Casañas A., Querol-Audí J., Guerra P., Pous J., Tanaka H., Tsukihara T., Verdaguer N., Fita I. New features of vault architecture and dynamics revealed by novel refinement using the deformable elastic network approach. (англ.)