Глюконеогенез
Глюконеогене́з — метаболічний шлях утворення глюкози з нецукрових вуглецевих субстратів, таких як піруват, молочна кислота, гліцерин і глюкогенні амінокислоти. Глюкоза займає центральну роль в метаболізмі всіх організмів, від бактерій до людини, вона є універсальним джерелом енергії та будівельним блоком для синтезу багатьох речовин. Деякі органи людини використовують глюкозу як єдиний або основний енергетичний субстрат, наприклад мозок, еритроцити, кора нирок, яєчка, ембріональні тканини. Один тільки головний мозок потребує близько 120 г глюкози в день, тобто більше половини всієї глюкози, що зберігається у формі глікогену у скелетних м'язах та печінці. В періоди між прийомами їжі, довшого голодування або інтенсивних фізичних навантажень запас глюкози може вичерпуватись, тому існує метаболічний шлях глюконеогенезу, що забезпечує її утворення із невуглеводневих попередників, таких як піруват та близькі три- або чотирикарбонові сполуки. Глюконеогенез є енергозатратним процесом.
Метаболічний шлях глюконеогенезу присутній у представників всіх основних груп живої природи: бактерій, архебактерій, рослин, грибів і тварин. Реакції глюконеогенезу однакові у всіх організмів у всіх тканинах, проте може відрізнятись його метаболічний контекст.
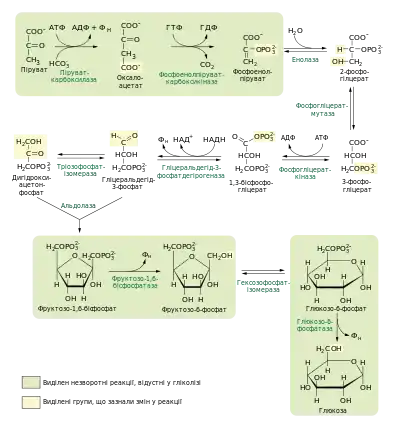
Глюконеогенез забезпечує синтез глюкози із пірувату, а гліколіз навпаки — розщеплення глюкози до пірувату, проте глюконеогенез не є оберненою копією гліколізу, хоча багато реакцій (сім із десяти) є спільними для обидвох шляхів. Три реакції гліколізу дуже екзергонічними (тобто мають велику негативну зміну вільної енергії) і незворотні у живих клітинах: перетворення глюкози до глюкозо-6-фосфату, перетворення фруктозо-6-фосфату до фруктозо-1,6-бісфосфату та перетворення фосфоенолпірувату (ФЕП) до пірувату (див. гліколіз). У глюконеогенезі є обхідні шляхи (шунти) для цих реакцій, які також мають велику негативну зміну вільної енергії. Таким чином обидва шляхи — і гліколіз, і глюконеогенез — є незворотніми у клітині.
Локалізація і значення
Глюконеогенез відбувається у клітинах бактерій, архебактерій, грибів, рослин і тварин. Як і гліколіз, майже всі перетворення глюконеогенезу локалізується у цитоплазмі, проте в еукаріот перша реакція цього шляху має місце у мітохондріях[1].
У тварин найважливішими попередниками глюкози є тривуглецеві сполуки, такі як піруват, лактат, гліцерол та деякі амінокислоти. У ссавців глюконеогенез найбільш інтенсивно протікає у печінці, а також деякою мірою у кірковому шарі нирок та епітелії тонкого кишківника[1]. За добу в організмі людини синтезується до 80 г глюкози[2]. Після фізичних навантажень молочна кислота утворена в скелетних м'язах переноситься кров'ю до печінки, де перетворюється у глюкозу, яка транспортується назад у м'язи і слугує там субстратом для синтезу глікогену. Цей метаболічний шлях отримав назву цикл Корі[1]. Глюконеогенез відіграє особливе значення під час голодування, так методом ізотопного мічення було показано що на 22 годину утримання від вживання їжі він забезпечує утворення 64% всієї глюкози в крові, а на 46 годину ця цифра наближається до 100%[3].
Глюконеогенез також інтенсивно відбувається у насінні, яке проростає, і є частиною шляху, що перетворює запасні ліпіди та білки у дисахариди (переважно сахарозу), які можуть транспортуватись у всі тканини молодої рослини. Також фотоавтотрофам глюконеогенез потрібний для перетворення первинних продуктів фотосинтезу до глюкози. Остання необхідна рослинам для синтезу клітинної стінки та як попередник нуклеотидів, коферментів та багатьох інших речовин[1].
Багато мікроорганізмів починають глюконеогенез із двокарбонових та трикарбонових сполук, наявних у середовищі, де вони живуть, таких як ацетат, лактат, пропіонат[1].
Реакції глюконеогенезу
Сім реакцій глюконеогенезу є оберненими до реакцій гліколізу. Енергетичний бар'єр трьох незворотних гліколітичних реакцій долається у глюконеогенезі обхідними шляхами, до них належать: синтез фосфоенолпірувату із пірувату, перетворення фруктозо-1,6-бісфосфату у фруктозо-6-фосфат та перетворення глюкозо-6-фосфату до глюкози[1]. Така організація протилежних метаболічних шляхів не тільки дозволяє їм обидвом бути термодинамічно вигідними за однакових умов, а й дає можливість для їх роздільної регуляції[4].
Синтез фосфоенолпірувату із пірувату
Остання реакція гліколізу — перетворення фосфоенолпірувату у піруват із одночасним фосфорилюванням АДФ — має велику негативну зміну вільної енергії і тому є незворотною. В глюконеогенезі протилежне перетворення (пірувату у фосфсфоенолпіруват) відбувається обхідним шляхом, що складається як мінімум із двох реакцій[5], і в еукаріот потребує ферментів як мітохондрій так і цитоплазми. Протікання цієї стадії відрізняється в залежності від того піруват чи лактат є попередником у синтезі глюкози[6].
Піруват спершу перетворюється до оксалоацетату вналслідок карбоксилювання піруваткарбоксилазою. Цей фермент використовує біотин як кофермент, реакція супроводжується гідролізом однієї молекули АТФ. Біотин виступає носієм бікарбонату, що попередньо активується шляхом утворення змішаного ангідриду (карбоксифосфату) внаслідок перенесення фосфатної групи із АТФ[7]. Рівняння реакції:
- Піруват + АТФ + HCO-3 → оксалоацетат + АДФ + Фн;
Реакція карбоксилювання необхідна для метаболічної активації пірувату[8][9].
Наступна реакція — одночасне декарбоксилювання та фосфорилювання оксалоацетату — каталізується ферментом фосфоенолпіруваткарбоксикіназою, що потребує присутності іонів Mg2+ та ГТФ як донора фосфатної групи. Продуктом цієї реакції є фосфоенолпіруват, вона зворотна за клітинних умов[8].
- Оксалоацетат + ГТФ → фосфоенолпіруват + ГДФ + CO2;
Сумарне рівняння процесу:
- Піруват + АТФ + ГТФ + HCO-3 → Фосфоенолпіруват + АДФ + ГДФ + Фн + CO2, ΔG0= 0,9 кДж/моль.
Таким чином для перетворення пірувату до фосфоенолпірувату необхідний гідроліз двох молекул нуклеотидтрифосфатів, тоді як протилежний процес у гліколізі дозволяє синтезувати тільки одну молекулу АТФ. Хоча стандартна зміна вільної енергії для сумарного процесу становить 0,9 кДж/моль, в реальних умовах завдяки дуже низькій концентрації фосфоенолпірувату ΔG = −25 кДж/моль, тобто перетворення є сильно екзергонічним і незворотним[8].
Човниковий транспорт оксалоацетату
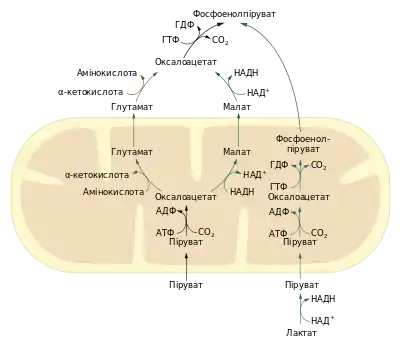
Утворення оксалоацетату є так званою анаплеротичною реакцією циклу трикарбонових кислот, тобто такою, що підтримує достатній рівень його метаболітів[10][11]. Тому, як і сам ЦТК, вона відбувається у матриксі мітохондрій, піруваткарбоксилаза є виключно мітохондріальним ферментом в еукаріот. Натомість локалізація ФЕП-карбоксикінази відрізняється у різних організмів: у печінці мишей та пацюків вона міститься тільки в цитозолі, у кролів та голубів — тільки в мітохондріях, а в людини та морських свинок приблизно порівну розподілена між двома компартментами[12]. Решта ж ферментів глюконеогенезу є цитозольними, таким чином для проходження цього метаболічного шляху оксалоацетат або фосфоенолпіруват повинні транспортуватись із мітохондрій у цитоплазму. Конкретний механізм транспорту залежить від організму та речовини, що виступає попередником у синтезі глюкози.
Якщо попередником є піруват, то використовується переважно малатний шлях транспорту. Піровиноградна кислота переноситься до матриксу мітохондрій або утворюється там із амінокислоти аланіну в реакції переамінування, тут відбувається карбоксилазна реакція. Утворений оксалоацетат не може бути транспортований до цитзолю, через те, що внутрішня мембрана мітохондрій не має для нього транспортера. Тому оксалоацетат відновлюється малатдегідрогеназою до малату за рахунок перенесення гідрид іона із НАДH. Попри те, що стандартна зміна вільної енергії для цієї реакції досить висока, за умов характерних для матриксу мітохондрій (зокрема високої концентрації оксалоацетату), вона є оборотною (ΔG~0). Утворений L-малат покидає мітохондрії за посередництва спеціального переносника і в цитоплазмі знову окиснюється до оксалоацетату. Останній перетворюється до ФЕП. Цей шлях забезпечує експорт у цитозоль не тільки оксалоацетату а й відновних еквівалентів НАДH, необхідних для протікання глюконеогензу (відновлення 1,3-бісфосфогіцерату до гліцеральдегід-3-фосфату). В цитоплазмі співвідношення НАДH/НАД+ становить близько 8×10−4 і є в сто тисяч раз меншим ніж у мітохондріях. Утворення малату у матриксі мітохондрій, його транспорт у цитоплазму і дегідрогенізація забезпечує баланс між утвореним та використаним НАДH у цитоплазмі під час глюконеогенезу[8].
Дещо відрізняється початок глюконеогенезу в тому випадку, коли субстратом для синтезу глюкози слугує лактат (утворений в еритроцитах або скелетних м'язах під час інтенсивних навантажень). В такому разі молочна кислота дегідрогенізується у цитоплазмі, ця реакція є джерелом НАДH, а отже нема потреби у перенесенні відновних еквівалентів у вигляді малату із мітохондрій. Утворений піруват транспортується до мітохондрій, де є субстратом для піруваткарбоксилази. Після цього оксалоацетат відразу ж у матриксі підлягає декарбоксилюванню та фосфорилюванню завдяки мітохондріальній фосфоенолпіруваткарбоксикіназі. Утворений фосфоенолпіруват покидає мітохондрії[13].
Існує ще один шлях, що не передбачає перенесення НАДH — аспартатний. У цьому випадку оксалоацетат у матриксі вступає в реакцію переамінування з амінокислотами катлізовану аспартатамінотрансферазою. Внаслідок цього він перетворюється в аспартат, який транспортується в цитозоль. Там знову відбувається переамінування за участі аспартатамінотрасферази, внаслідок чого утворюється оксалоацетат. Цей шлях також використовується тоді, коли попередником у глюконеогенезі є молочна кислота, зокрема організмами що не містять мітохондріальної ФЕП-каброксикінази[14].
Фосфорилазні реакції глюконеогенезу
Дві інші незворотні стадії гліколізу — кіназні реакції: фосфорилювання фруктозо-6-фосфату та глюкози із використанням АТФ. Обернені реакції вимагали би перенесення фосфатної групи із фосфорильованих моносахаридів назад на АДФ, проте в глюконеогенезі цього не відбувається, відповідні перетворення натомість каталізуються іншими ензимами — фосфатазами (фруктозо-1,6-бісфосфатазою (ФБФ-1) та глюкозо-6-фосфатазою). Фосфатазні реакції — це простий гідроліз, продуктом якого є фосфатна кислота[13][9]:
- Фруктозо-1,6-бісфосфат + H2O → фруктозо-6-фосфат + Фн;
- Глюкозо-6-фосфат + H2O → глюкоза + Фн.
Обидва ферменти є магній-залежними. Глюкозо-6-фосфатаза відсутня у більшості тканин, тому глюконеогенез в них завершується формуванням глюкозо-6-фосфату, який може бути використаний для синтезу глікогену або участі в інших метаболічних шляхах. Такі тканини не здатні поповнювати рівень глюкози в крові, оскільки глюкозо-6-фосфат не може транспортуватись плазматичною мембраною[9]. Глюкозо-6-фосфатаза присутня у гепатоцитах, та, в меншій мірі, в клітинах печінки та епітелію тонкого кишківника[13][15]. Локалізується вона у порожнині ендоплазматичного ретикулуму, куди спеціальним переносником транспортується глюкозо-6-фосфат, а пізніше іншим транспортним білком викачується глюкоза і фосфат[9].
Енергетичні затрати глюконеогенезу
Формування глюкози із пірувату є термодинамічно невигідним процесом, тому воно повинне бути спряжене із екзергонічними реакціями, а саме гідролізом нуклеотидтрифосфатів[16]. Сумарне рівняння глюконеогенезу, у випадку, коли вихідною речовиною виступає піруват, має такий вигляд:
- 2 Піруват + 4АТФ + 2ГТФ + 2НАДH(H+) + 4H2O → глюкоза + 4АДФ + 2ГТФ + 6Фн + 2НАД+;
Отже для утворення однієї молекули глюкози необхідна енергія шести високоенргетичних фосфатних груп (чотирьох від АТФ та двох від ГТФ). Також в цьому процесі використовуються дві молекули НАДH для відновлення 1,3-бісфосфогліцерату.
Для порівняння сумарне рівняння гліколізу:
- Глюкоза + 2АДФ + 2Фр + НАД+ → 2 піруват + 2АТФ + 2H2O + НАДH(H+);
Очевидно, що глюконеогенез не є просто оберненим до гліколізу, оскільки в такому разі для його проходження вистачало би всього двох молекул АТФ. Глюконеогенез відносно енергетично «дорогий» метаболічний шлях, багато із енергії потрібно для забезпечення його незворотності. За клітинних умов сумарна зміна вільної енергії у процесі гліколізу становить близько −63 кДж/моль, а в глюконеогенезі — −16 кДж/моль[17].
Попередники у синтезі глюкози
| Глюкогенні амінокислоти[17] | |
|---|---|
| Аланін | Піруват |
| Цистеїн | |
| Гліцин | |
| Серин | |
| Треонін | |
| Триптофан | |
| Аргінін | α-Кетоглутарат |
| Глутамат | |
| Глутамін | |
| Гістидин | |
| Пролін | |
| Ізолейцин | Сукциніл-КоА |
| Метіонін | |
| Треонін | |
| Валін | |
| Фенілаланін | Фумарат |
| Тирозин | |
| Аспарагін | Оксалоацетат |
| Аспартат | |
Піруват і проміжні продукти ЦТК
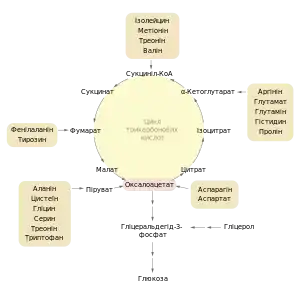
Описаний метаболічний шлях глюконеогенезу може використовуватись для біосинтезу глюкози не тільки зі пірувату та лактату, а також і багатьох інших речовин, зокрема проміжних продуктів циклу трикарбонових кислот (ЦТК). Такі сполуки як цитрат, ізоцитрат, α-кетоглутарат, сукциніл-КоА, сукцинат, фумарат та малат перетворюються в ході ЦТК до оксалоацетату, а отже можуть бути субстратами для глюконеогенезу[17][3].
Карбонові скелети більшості амінокислот також можуть бути перетворені до пірувату або проміжних продуктів ЦТК (див. таблицю зліва) і виступати попередниками глюкози, вони називаються глюкогенними. Серед 20 стандартних амінокислот тільки дві: лейцин та лізин не можуть використовуватись для синтезу глюкози[17][3].
Серед глюкогенних амінокислот найбільше значення для глюконеогенезу мають аланін та глутамін, оскільки вони виступають основними переносниками аміногруп від різних органів до печінки. У мітохондріях гепатоцитів від них відщеплюються аміногрупи, а карбонові скелети використовуються на біосинтез глюкози[17].
Гліцерол
Попередником у синтезі глюкози також може виступати продукт гідролізу нейтральних жирів гілцерол. Для цього у клітинах печінки він фосфорилюється гліцеролкіназою, після чого відбувається окиснення другого атома Карбону і утворюється гліцеральдегід-3-фосфат, який може вступати в глюконеогенез. Хоча гліцеролфосфат є важливим попередником у синтезі тригліцеридів в адипоцитах, ці клітини не мають гліцеролкінази. Тому вони використовують для синтезу цієї речовини скорочений варіант глюконеогенезу: гілцеронеогенез, який включає перетворення пірувату до дигідроксиацетонфосфату із його подальшим відновленням до гліцеролфосфату[17].
Жирні кислоти
Більшість жирних кислот (як і дві неглюкогенні амінокислоти лейцин та лізин) розщеплюються до ацетил-КоА, який в організмі тварин не може бути використаний для глюконеогензу через відсутність метаболічного шляху перетворення ацетил-КоА у піруват або оксалоацетат[17][3]. Проте у рослин, деяких грибів (зокрема дріжджів), водоростей, найпростіших та бактерій наявний модифікований варіант ЦТК гліоксилатний цикл, що дозволяє синтезувати оксалоацетат із ацетил-КоА[18]. Таким чином ці організми можуть використовувати жирні кислоти для синтезу глюкози. У рослин цей шлях важливий під час проростання насіння, до того часу як у молодої рослини з'являться листки і почнеться повноцінний фотосинтез, вона повинна покладатись на запасні жири як джерело енергії і будівельних блоків, зокрема для синтезу клітинних стінок[17].
Регуляція глюконеогенезу
Якби гліколіз та глюконеогенез могли протікати одночасно з високою інтенсивністю в клітині, результатом було би даремне споживання енергії і перетворення її у тепло. Наприклад фосфофруктокіназна та фруктозо-1,6-фосфатазна реакції:
- Фруктозо-6-фосфат + АТФ → фруктозо-1,6-бісфосфат + АДФ;
- Фруктозо-1,6-бісфосфат + H2O → фруктозо-6-фосфат + Фн;
давали би в сумі тільки гідроліз АТФ (відбувається так званий субстратний цикл):
- АТФ + H2O → АДФ+ Фн.
Тому ці два шляхи реципрокно регулюються алостерично, шляхом ковалентної модифікації ферментів та регуляції їхнього синтезу. На швидкість глюконеогенезу також впливає доступність субстратів. Загалом, коли клітині потрібна енергія, в ній більш активно відбувається гліколіз, а коли енергії є в надлишку, то переважатиме глюконеогенез[17][16].
Регуляція піруваткарбоксилази
Піруваткарбоксилаза є першим регуляторним ферментом глюконеогенезу. Для функціонування вона потребує приєднання алостеричного активатора ацетил-КоА, високий рівень якого свідчить про достатній запас жирних кислот, які можуть бути окиснені з метою отримання енергії[7]. Проте продукт піруваткарбоксилазної реакції — оксалоацетат — використовуватиметься на поповнення циклу трикарбонових кислот, а не на глюконеогенез, якщо тільки ЦТК не інгібуватиметься високими рівнями АТФ або НАДH[19]. Негативним модулятором піруваткарбоксилази є АДФ[20].
Регуляція ФЕП-карбоксикінази
ФЕП-карбоксикіназа каталізує перший комітований крок глюконеогенезу (тобто такий, що однозначно визначає метаболізм певної сполуки по цьому шляху). У ссавців його регуляція відбувається переважно на транскрипційному рівні у відповідь на зміну дієти та рівня гормонів. Зокрема, глюкагон, глюкокортикоїди та тироїдні гормони стимулюють синтез ФЕП-каброксикінази, в той час як інсулін пригнічує її. Наприклад, під час голодування у крові підвищується рівень глюкагону, що викликає синтез цАМФ у гепатоцитах. Останній зв'язується із транскрипційним фактором CREB (англ. CRE-binding protein, CRE-зв'язуючий білок), а останній приєднується до регуляторної ділянки CRE (англ. cAMP response element, цАМФ-чутливий елемент) у промоторі гену ФЕП-карбоксикінази, активуючи експресію останнього[21].
Регуляція фруктозо-1,6-бісфосфатази

Останній регуляторний фермент глюконеогенезу інгібується АМФ, високий рівень якого свідчить про вичерпання запасів АТФ[20]. У гепатоцитах його активність прив'язана до рівня глюкози в крові завдяки сигнальній молекулі фруктозо-2,6-бісфосфату, яка одночасно виступає алостеричним інгібітором фруктозо-1,6-бісфосфатази та алостеричним активатором відповідного ферменту гліколізу — фосфофруктокінази. Концентрація фруктозо-2,6-бісфосфату залежить від швидкості її утворення із фруктозо-6-фосфату фосфофруктокіназою-2 (ФФК-2) та гідролізу фруктозо-2,6-бісфосфатазою (ФБФаза-2). ФФК-2 та ФБФаза-2 — це дві різні активності одного біфункціонального ферменту, який «перемикається» шляхом фосфорилювання[17][22].
У випадку, коли рівень глюкагону в крові високий, він стимулює у гепатоцитах цАМФ-залежний сигнальний шлях, що призводить до фосфорилювання біфункціонального ферменту протеїнкіназою А. Фосфорильована форма цього білка функціонує як ФБФаза-2 і гідролізує фруктозо-2,6-бісфосфат, внаслідок чого відбувається активація фруктозо-1,6-бісфосфатази і пригнічення фосфофруктокінази-1. Отже глюконеогенез відбувається інтенсивніше ніж гліколіз. Інсулін викликає протилежну відповідь: дефосфорилювання біфункціонального ферменту, збільшення концентрації фруктозо-2,6-бісфосфату, активацію ФФК-1 та пригнічення ФБФази-1[23][22].
Примітки
- Nelson et al, 2008, с. 552.
- Губський, 2007, с. 212.
- Voet et al, 2011, с. 871.
- Voet et al, 2011, с. 872.
- Prescott, 2002, с. 209.
- Nelson et al, 2008, с. 553.
- Nelson et al, 2008, с. 554.
- Nelson et al, 2008, с. 555.
- Berg et al, 2007, с. 463.
- Губський, 2007, с. 213.
- Voet et al, 2011, с. 873.
- Voet et al, 2011, с. 876.
- Nelson et al, 2008, с. 556.
- Voet et al, 2011, с. 877.
- Voet et al, 2011, с. 887.
- Berg et al, 2007, с. 464.
- Nelson et al, 2008, с. 557.
- Prescott, 2002, с. 216.
- Voet et al, 2011, с. 874.
- Berg et al, 2007, с. 465.
- Voet et al, 2011, с. 879.
- Berg et al, 2007, с. 466.
- Nelson et al, 2008, с. 558.
Джерела
- Berg JM, Tymoczko JL, Stryer L (2007). Biochemistry (вид. 6th). W.H. Freeman and Company. ISBN 0-7167-8724-5.
- Nelson D.L., Cox M.M. (2008). Lehninger Principles of Biochemistry (вид. 5th). W. H. Freeman. ISBN 978-0-7167-7108-1.
- Prescott L.M. (2002). Microbiology (вид. 5th). McGraw−Hill. ISBN 0-07-282905-2.
- Voet D., Voet J.G. (2011). Biochemistry (вид. 4th). Wiley. с. 487—496. ISBN 978-0470-57095-1.
- Губський Ю.І. (2007). Біологічна хімія. Київ-Вінниця: Нова книга. с. 191. ISBN 978-966-382-017-0.
Посилання
- ГЛЮКОНЕОГЕНЕЗ //Фармацевтична енциклопедія
- Глюконеогенез // Універсальний словник-енциклопедія. — 4-те вид. — К. : Тека, 2006.
