Мікроглія
Мікроглія — тип нейроглії (гліальних клітин), розташованої по всьому головному та спинному мозку.[1] Мікроглія становить 10–15 % усіх клітин мозку.[2] Як макрофаги-резиденти, вони діють як перша та основна форма активного імунного захисту в центральній нервовій системі (ЦНС).[3] Мікроглія (та інші нейроглії, включаючи астроцити) поширюються у великих регіонах, що не перекриваються, по всій ЦНС.[4][5] Мікроглія є ключовими клітинами в загальному обслуговуванні мозку — вони постійно «очищують» ЦНС від сенільних бляшок, пошкоджених або непотрібних нейронів і синапсів та інфекційних агентів.[6] Оскільки ці процеси повинні бути ефективними для запобігання потенційно смертельних пошкоджень, мікроглія надзвичайно чутлива навіть до невеликих патологічних змін у ЦНС.[7] Ця чутливість частково досягається наявністю унікальних калієвих каналів, які реагують навіть на незначні зміни позаклітинного калію. Останні дані свідчать, що мікроглія також є ключовим гравцем у підтримці нормальних функцій мозку в здорових умовах.[8] Мікроглія також постійно контролює функції нейронів за допомогою прямих соматичних контактів і надає нейропротекторні ефекти, коли це необхідно.[9]
Структури ендотеліальних клітин, відомі як гематоенцефалічний бар'єр, або ГЕБ, не допускають патогени з крові безпосередньо до головного та спинного мозку, з яких складається ЦНС. ГЕБ запобігає потраплянню більшості інфекцій до вразливої нервової тканини. У тому випадку, коли збудники інфекції потрапляють безпосередньо в мозок або перетинають гематоенцефалічний бар'єр, мікрогліальні клітини повинні швидко реагувати щоб зменшити запалення та знищити збудники інфекції, перш ніж вони пошкодять чутливу нервову тканину. Через недостатність антитіл (небагато антитіл досить малі, щоб пройти ГЕБ), мікроглія повинна мати можливість розпізнавати сторонні об'єкти, поглинати їх та діяти як антигенопрезентувальні клітини, які активують Т-клітини .
Форми
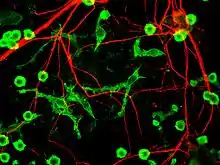
_in_rat_cerebellar_molecular_layer.jpg.webp)
Мікрогліальні клітини надзвичайно пластичні та зазнають різноманітних структурних змін залежно від місця та системних потреб. Цей рівень пластичності необхідний для виконання широкого спектру їх функцій. Здатність до трансформації відрізняє мікроглію від макрофагів, які повинні регулярно замінюватись, і забезпечує здатність мікроглії захищати ЦНС в надзвичайно короткі терміни, не викликаючи імунологічних порушень.[6] Мікроглія приймає певну форму, або фенотип, у відповідь на місцеві умови та виявлені ними хімічні сигнали.
Генетика сенсом
Сенсоми мікроглії — відносно нова біологічна концепція, яка, мабуть, відіграє велику роль у розвитку нервової системи та її дегенерації. Сенсоми належать до унікальній групи транскриптів білка, що використовуються для розпізнавання лігандів та мікробів. Іншими словами, сенсоми — це гени, необхідні для синтезування білків, що використовуються для розпізнавання молекул в організмі. Сенсоми можна проаналізувати за допомогою різних методів, включаючи qPCR, РНК-послідовності, аналіз мікрочипів та пряме секвенування РНК. Гени, включені в сенсомний код рецепторів та трансмембранних білків на плазматичній мембрані, сильніше експресуються в мікроглії порівняно з нейронами. Цей код не включає білки, які секретуються, або трансмембранні білки, специфічні для мембранних органел, таких як ядро, мітохондрії та ендоплазматичний ретикулум.[10] Ідентифіковано багато сенсомних генів, які кодують рецептори розпізнавання паттернів, однак серед них існує велика різноманітність. Мікроглія має сенсом подібний до інших макрофагів, однак вона має 22 унікальних гени, 16 з яких використовуються для взаємодії з ендогенними лігандами. Ці відмінності створюють унікальний мікрогліальний біомаркер, що включає понад 40 генів, включаючи P2ry12 та HEXB. DAP12, мабуть, відіграє важливу роль у взаємодії сенсорних білків, виступаючи в якості сигнального адаптера та регуляторного білка.
Регуляція генів у сенсомі має мати можливість змінюватися, щоб реагувати на потенційну шкоду. Мікроглія може взяти на себе роль нейропротектора або нейротоксина, щоб протистояти цим небезпекам.[11] З цих причин існує припущення, що сенсом може відігравати роль у нейродегенерації. Сенсомічні гени, які активуються при старінні, в основному беруть участь у розпізнаванні інфекційних мікробних лігандів, тоді як ті, активність яких знижується, в основному беруть участь у розпізаванні ендогенних лігандів.[10] Цей аналіз свідчить про специфічну для глії регуляцію, що сприяє нейропротекції під час природної нейродегенерації. Це контрастує зі здвигом убік її нейротоксичного впливу, який спостерігається при нейродегенеративних захворюваннях.
Сенсом також може зіграти свою роль у розвитку нервової системи. Інфекція головного мозку на початку життя призводить до формування мікроглії, гіперчутливої до подальших імунних подразників. При впливі інфекції, спостерігається активація сенсомальних генів, що беруть участь в нейрозапаленні і гальмування генів, які пов'язані з нейропластичністю.[12] Здатність сенсому змінювати розвиток нервової тканини, однак, може використовуватись у боротьбі із захворюваннями. Делеція CX3CL1, високоекспресованого сенсомного гена, в моделях синдрому Ретта на гризунах призвело до поліпшення стану здоров'я та збільшення тривалості життя.[13] Зниження активності Cx3cr1 у людей без синдрому Ретта пов'язане із симптомами, подібними до шизофренії.[14] Це свідчить про те, що сенсом не тільки відіграє роль у різних порушеннях розвитку, але також вимагає ретельної регуляції, щоб підтримувати безхворобний стан.
Гілляста мікроглія
Ця форма мікрогліальних клітин зазвичай зустрічається в певних місцях у всьому мозку та спинному мозку за відсутності чужорідного матеріалу або відмираючих клітин. Ця «мікроглія у стані спокою» складається з довгих відгалужень і малого клітинного тіла. На відміну від амебоїдних форм мікроглії, тіло гіллястих форм залишається на місці, тоді як її гілки постійно рухаються та «оглядають» навколишній просір. Гілки дуже чутливі до незначних змін у фізіологічному стані і потребують дуже специфічних умов культури для спостереження in vitro.
На відміну від активованої або амебоїдної мікроглії, гілляста мікроглія не фагоцитує клітини і виділяє менше імуномолекул (включаючи білки ГКГС класу I/ІІ). Мікроглія в цьому стані здатна шукати та виявляти імунні загрози, зберігаючи гомеостаз у ЦНС.[15][16][17] Незважаючи на те, що це вважається станом спокою, мікроглія в такій формі все ще надзвичайно активно займається хімічним «обстеженням» навколишнього середовища. Гіллясту мікроглію можна перетворити в активовану форму у будь-який час у відповідь на травму чи загрозу.
Реактивна (активована) мікроглія
Хоча існує історично часто вживаний термін «активована мікроглія», він повинен бути замінений на «реактивна мікроглія».[18] Дійсно, очевидно мікроглія у стані спокою не позбавлена активних функцій, і термін «активація» вводить в оману, оскільки має тенденцію вказувати на поляризацію клітинної реактивності «все або нічого». Для візуалізації цих клітин часто використовується маркер Iba1, який активується в реактивній мікроглії.[19]
Нефагоцитуюча мікроглія
Цей стан насправді є проміжною формою, оскільки мікроглія переходить із гіллястої форми до повністю активної фагоцитуючої форми. Мікроглія може активуватися різними факторами, включаючи: прозапальні цитокіни, фактори некрозу клітин, ліпополісахарид та зміни позаклітинного рівня калію (що свідчить про розрив мембрани клітин). Після активації клітини зазнають декількох основних морфологічних змін, включаючи потовщення та втягнення гілок, поглинання білків ГКГС класу I/II, експресію імуномолекул, секрецію цитотоксичних факторів, секрецію рекрутингових молекул та секрецію прозапальних сигнальних молекул (в результаті в каскаді сигналів про запалення). Активована нефагоцитуюча мікроглія, як правило, виглядає як «кущасті», «паличкоподібні» або невеликі амебоїди, залежно від того, наскільки далеко вони знаходяться на шляху свого перетворення. Крім того, мікроглія швидко розмножується. З суцільно морфологічної точки зору, варіація форми мікроглії при її перетворенні пов'язана зі зміною морфологічної складності і може бути визначена кількісно за допомогою методів фрактального аналізу, які виявилися чутливими до навіть тонких, візуально не помітних змін, пов'язаних з різною морфологією в різних патологічних станах.[6][15][16][20]
Фагоцитуюча мікроглія
Активована фагоцитуюча мікроглія є максимально імунно-реагуючою формою. Ці клітини, як правило, набувають великої, амебоїдної форми, хоча спостерігалися деякі варіації. На додаток до функцій нефагоцитуючої мікроглії (презентація антигену, цитотоксичність та активація запалення), вона також здатна фагоцитувати чужорідні агенти та презентувати отримані імуномолекули для активації Т-клітин. Фагоцитуюча мікроглія подорожує до місця пошкодження, поглинає чужорідний матеріал та виділяє прозапальні фактори, для сприяння розмноженню більшої кількості клітин і також розмножується сама. Активована фагоцитуюча мікроглія також взаємодіє з астроцитами та нервовими клітинами, щоб якомога швидше реагувати на будь-яку інфекцію або запалення з мінімальним пошкодженням здорових клітин мозку.[6][15]
Амебоїдна мікроглія
Ця форма дозволяє мікроглії вільно пересуватися по нервовій тканині, завдяки чому вона виконує свою роль клітини-поглинача. Амебоїдна мікроглія здатна фагоцитувати уламки, але не виконує антигенопрезентувальну та запальну роль, притаманну активованій мікроглії . Амебоїдна мікроглія особливо превалює під час розвитку та ремоделювання мозку, коли є велика кількість позаклітинного дебрісу та апоптотичних клітин. Ця форма мікроглії присутня переважно в перинатальних ділянках білої речовини в мозолистому тілі, що описується як «фонтани мікроглії».[6][16][21]
Гіттерні клітини
Гіттерні клітини є результатом фагоцитозу інфекційного матеріалу або клітинного дебрісу («сміття») гліальними клітинами. Зрештою, після поглинання певної кількості матеріалу, фагоцитарна мікрогліальна клітина стає нездатною для подальшого фагоцитозу. Отримана клітинна маса відома як «гранулярні тільця», названі за їх «зернистий» вигляд. Дивлячись на тканини, забарвлені для виявлення гліттерних клітин, патологоанатоми можуть візуалізувати ділянки загоєння після інфекції.[22]
Периваскулярна мікроглія
На відміну від інших типів мікроглії, згаданих вище, назва «периваскулярна мікроглія» стосується місця розташування клітин, а не їх форми/функції. Периваскулярна мікроглія розташовується в основному в межах базальної пластинки. Вона виконує нормальні мікрогліальні функції, але, на відміну від нормальної мікроглії, вона регулярно замінюється клітинами-попередниками з кісткового мозку та експресує антигени ГКГС класу II незалежно від їх оточення. Периваскулярна мікроглія також сильно реагує на антигени диференціації макрофагів.[6] Встановлено, що ця мікроглія має важливе значення для відновлення судинних стінок, що показали експерименти та спостереження Ріттера при ішемічній ретинопатії. Периваскулярна мікроглія сприяє проліферації ендотеліальних клітин, дозволяючи формування нових та відновлювання пошкоджених судин. В ході цих процесів, рекрутування мієлоїдних клітин і диференціація їх у мікроглію сильно прискорюються для виконання цих завдань.[23]
Юкставаскулярна мікроглія
Як і периваскулярна, юкставаскулярна мікроглія розрізняється головним чином за місцем її розташування. Юкставаскулярна мікроглія має безпосередній контакт з базальною пластинкою судин, але в самих стінках не виявляється. Як і периваскулярна мікроглія, вона експресує білки ГКГС класу II навіть при низькому рівні запальних цитокінів. На відміну від периваскулярних клітин, але подібно до резидентної мікроглії, юкставаскулярна мікроглія не демонструє швидкого обороту або заміщення мієлоїдними клітинами-попередниками на регулярній основі.[6]
Функції
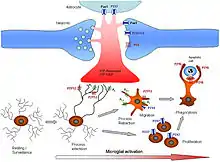
Клітини мікроглії виконують різноманітні завдання в ЦНС, в основному пов'язані як з імунною реакцією, так і з підтримкою гомеостазу. Нижче наведено деякі основні відомі функції, які виконують ці клітини.
Скавенджинг
Окрім надчутливості до малих змін у своєму середовищі, кожна мікрогліальна клітина також регулярно фізично «обстежує» свій домен. Ця дія здійснюється в амебоїдному та спокійному станах. Якщо мікрогліальна клітина, переміщуючись по заданій ділянці, «знайде» сторонній матеріал, пошкоджені клітини, апоптовані клітини, нейрофібрилярні клубки, фрагменти ДНК або бляшки, вона активується і фагоцитує матеріал або клітину. Таким чином мікрогліальні клітини також виступають в ролі «хатніх помічників», прибираючи випадкові клітинні залишки.[15] Під час розвитку в період формування міжнейронних зв'язків мікрогліальні клітини відіграють велику роль в регулюванні кількості клітин-попередників нейронів та видаленні апоптованих нейронів. Існують також докази того, що мікроглія може вдосконалити синаптичні схеми, поглинаючи та усуваючи синапси.[24] Після розвитку головного мозку більшість мертвих або апоптованих клітин знаходиться в корі та підкірковій білій речовині. Це може пояснити, чому більшість амебоїдних мікрогліальних клітин знаходяться в «фонтанах мікроглії» в корі головного мозку.[21]
Фагоцитоз
Основна роль мікроглії — фагоцитоз, включає поглинання різних матеріалів. Поглинені матеріали, як правило, складаються з клітинного дебрісу, ліпідів, апоптованих клітин у відсутності запалення, інвазивних вірусів, бактерій та інших чужорідних матеріалів при запаленні. Після того, як мікрогліальна клітина «наповнюється», вона припиняє фагоцитарну активність і перетворюється на відносно нереактивну гіттерну клітину .[25]
Позаклітинна сигналізація
Значна роль мікроглії — підтримка гомеостазу в неінфікованих ділянках та сприяння запаленню в інфікованій або пошкодженій тканині. Мікроглія досягає цього за допомогою надскладного набору позаклітинних сигнальних молекул, які дозволяють їм спілкуватися з іншою мікроглією, астроцитами, нервами, Т-клітинами та мієлоїдними клітинами-попередниками. Як згадувалося вище, цитокін IFN-γ може бути використаний для активації мікрогліальних клітин. Крім того, після активації за допомогою IFN-γ, мікроглія вивільняє ще більше IFN-γ у позаклітинний простір. Це активує ще більше мікроглії і запускає каскад активації, що швидко активує всю сусідню мікроглію. Мікроглія продукує ФНО-α, який призводить до апоптозу нервової тканини та посилює запалення. IL-8 сприяє зростанню та диференціації В-клітин, що допомагає мікроглії у боротьбі з інфекцією. Інший цитокін, IL-1, інгібує цитокіни IL-10 і TGF-β, які пригнічують презентацію антигену та прозапальну сигналізацію. Додаткові дендритні клітини та Т-клітини рекрутуються до місця пошкодження завдяки хемотаксичним молекулам, таким як MDC, IL-8 та MIP-3β, які також продукуються мікроглією. Нарешті, PGE 2 та інші простаноїди запобігають хронічному запаленню, пригнічуючи мікрогліальну протизапальну реакцію та знижуючи регуляцію Th1 (Т-хелперної клітини).[15]
Антигенопрезентація
Як вже згадувалося вище, резидентна неактивована мікроглія є слабкими антигенопрезентувальними клітинами через відсутність у них білків ГКГС класу I/II. Після активації вони швидко виробляють ці білки та швидко стають ефективними антигенопрезентерами. У деяких випадках антигенопрезентувальну функцію мікроглії також можна активувати за допомогою IFN-γ , але вона не функціонує настільки ефективно, як при експресії білків ГКГС класу I/II. Під час запалення Т-клітини перетинають ГЕБ завдяки спеціалізованим поверхневим маркерам, а потім безпосередньо зв'язуються з мікроглією, для отримання від них антигенів . Після того, як вони їх отримали, Т-клітини виконують різноманітні ролі, включаючи прозапальне рекрутування клітин, формування клітин пам'яті, секрецію цитотоксичних сполук та прямі атаки на плазматичні мембрани чужорідних клітин.[6][15]
Цитотоксичність
Крім того, що мікроглія здатна знищувати інфекційні організми шляхом контакту «клітина до клітини» за допомогою фагоцитозу, вона також може виділяти різноманітні цитотоксичні речовини.[26] Культура мікроглії виділяє велику кількість перекису водню та оксиду азоту в процесі, відомому як «респіраторний вибух». Ці обидві речовини можуть безпосередньо пошкоджувати клітини та призвести до загибелі нейронів. Протеази, що виділяються мікроглією, катаболізують специфічні білки, викликаючи пряме ушкодження клітин, тоді як цитокіни, такі як ІЛ-1, сприяють демієлінізації нейрональних аксонів. Нарешті, мікроглія може пошкоджувати нейрони за допомогою процесів, опосередкованих NMDA-рецепторами, шляхом виділення глутамату, аспартату та хінолінової кислоти . Цитотоксична секреція спрямована на знищення інфікованих нейронів, вірусів та бактерій, але також може спричинити велику кількість побічних нервових пошкоджень. В результаті хронічна запальна реакція може призвести до масштабних нервових пошкоджень, оскільки мікроглія руйнує мозок, намагаючись знищити проникаючу інфекцію.[6] Едаравон, поглинач радикалів, «вимикає» окислювальну нейротоксичність, викликану активованою мікроглією.[27]
Синаптичне оголення
У явищі, вперше поміченому при ураженні хребта Блінзінгером та Кройцбергом у 1968 році, мікроглія після запалення видаляє гілки з нервів біля пошкодженої тканини. Це сприяє відродженню та повторному формуванню (меппінг) пошкоджених нейронних мереж.[6]
Підтримка репарації
Після запалення мікроглія проходить кілька етапів, які сприяють відновленню нервової тканини. Вони включають синаптичне оголення, секрецію протизапальних цитокінів, рекрутинг нейронів та астроцитів у пошкоджену ділянку та утворення гіттерних клітин. Без мікроглії відновлення та повторне формування нейронних мереж у резидентних зонах ЦНС було б значно повільнішим і майже неможливим у багатьох судинних системах, що оточують мозок та очі.[6][23] Останні дослідження підтвердили, що мікроглія постійно контролює функції нейронів за допомогою спеціалізованих соматичних контактів і відчуває «самопочуття» нервових клітин. За допомогою цих міжклітинних комунікацій мікроглія здатна забезпечувати потужні нейропротекторні ефекти, значно сприяючи відновленню мозку після травми.[9]
Розвиток
Довгий час вважалося, що мікрогліальні клітини диференціюються в кістковому мозку з гемопоетичних стовбурових клітин-попередників усіх клітин крові. Однак останні дослідження показують, що мікроглія виникає в жовтковому мішку під час надзвичайно обмеженого ембріонального періоду і заселяє мезенхіму мозку. Крім того, вона постійно оновлюється і зберігається протягом усього життя без поповнення з периферійних моноцитарних попередників.[28]
Моноцити також можуть диференціюватися в мієлоїдні дендритні клітини та макрофаги в периферичних системах. Як і макрофаги в решті частин тіла, мікроглія використовує фагоцитарні та цитотоксичні механізми для руйнування чужорідних речовин. Мікроглія та макрофаги сприяють імунній відповіді, діючи як антигенопрезентувальні клітини, а також сприяючи запаленню та гомеостатичним механізмам в організмі шляхом секреції цитокінів та інших сигнальних молекул.[29]
У своїй неактивній формі мікроглія не має білків ГКГС класу I/ІІ, цитокінів IFN-γ, антигенів CD45 та багатьох інших поверхневих рецепторів, необхідних для виконання антигенопрезентувальні, фагоцитуючих та цитотоксичних ролей, які притаманні нормальним макрофагам. Мікроглія також відрізняється від макрофагів тим, що вона набагато щільніше регулюються у просторі та часі, для забезпечення точної імунної відповіді.[15]
Ще однією відмінністю між мікроглією та іншими клітинами, які походять з мієлоїдних клітин-попередників, є швидкість їх заміни. Макрофаги та дендритні клітини постійно використовуються і замінюються мієлоїдними клітинами-попередниками, які диференціюються у необхідний тип. Через ГЕБ організму досить складно постійно замінювати мікроглію. Тому замість того, щоб постійно замінюватись мієлоїдними клітинами-попередниками, мікроглія зберігає свій статус-кво, перебуваючи у стані спокою, а потім, після активації, вона швидко розмножується, щоб зберегти свою кількість. Дослідження кістково-мозкових химер показали, однак, що у випадках надзвичайно сильної інфекції ГЕБ ослаблюється, й мікроглія замінюється гематогенними клітинами, що походять з кісткового мозку, а саме мієлоїдними клітинами-попередниками та макрофагами. Після згасання інфекції бар'єр між периферійною та центральною системами відновлюється, і тільки сама мікроглія присутня у мозку протягом періоду відновлення.[30]
Старіння
На травму мікроглія реагує спалахом мітотичної активності; після цього спостерігається апоптоз із зменшенням кількості клітин до вихідного рівня.[31] Активація мікроглії навантажує анаболічний і катаболічний механізми клітин, змушуючи активовану мікроглію вмирати раніше, ніж неактивовані клітини.[31] Щоб компенсувати втрату мікроглії з плином часу, активується її мітоз, а клітини-попередники з кісткового мозку мігрують у головний мозок через його оболони та судини.[31]
Накопичення малих пошкоджень нейронів, що відбувається під час нормального старіння, може спричинити перетворення мікроглії в збільшені та активовані клітини.[32] Ця хронічна, вікова активація мікроглії та експресія IL-1 може з настанням віку сприяти збільшенню ризику хвороби Альцгеймера, сприяючи формуванню невритичних бляшок у прихильних до цього пацієнтів.[32] Пошкодження ДНК може сприяти віковій активації мікроглії. Іншим фактором може бути накопичення кінцевих продуктів глибокого глюкозування, які накопичуються зі старінням.[32] Ці білки дуже стійкі до протеолітичних процесів і сприяють крос-лінкінку («зшиванню») білків.[32]
Дослідження виявили у людини дистрофічну (внаслідок дефекту розвитку) мікроглію. «Ці клітини характеризуються аномаліями своєї цитоплазматичної структури, такими як відсутність розгалужень, атрофія, фрагментація або надзвичайна звивистість відростків, які часто мають сфероїдальні або цибулинні набрякання».[31] Частота дистрофічної мікроглії зростає зі старінням.[31] У дослідженнях пріонових хвороб, шизофренії та хвороби Альцгеймера повідомлялося про дегенерацію та загибель мікроглії, що свідчить про те, що її ушкодження може бути пов'язане з нейродегенеративними захворюваннями.[31] Слабким місцем цієї теорії є той факт, що відрізнити «активовану» та «дистрофічну» мікроглію в мозку людини важко.[31]
У мишей було показано, що блокада CD22 відновлює гомеостатичний мікрогліальний фагоцитоз у старіючих мізках.[33]
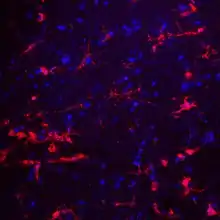
Клінічне значення
Мікроглія — це первинні імунні клітини центральної нервової системи, подібні до периферичних макрофагів. Вона реагує на патогени та травми, змінюючи морфологію та мігруючи до місця інфекції/травми, де знищує патогенні мікроорганізми та видаляє пошкоджені клітини. В рамках своєї реакції вона виділяє цитокіни, хемокіни, простагландини та активні форми кисню, які допомагають спрямувати імунну відповідь. Крім того, вона відіграє важливу роль у завершенні запальної реакції шляхом вироблення протизапальних цитокінів. Мікроглія також широко вивчалася на предмет їх шкідливої ролі при нейродегенеративних захворюваннях, таких як хвороба Альцгеймера, хвороба Паркінсона, розсіяний склероз, а також серцеві захворювання, глаукома, вірусні та бактеріальні інфекції. Накопичуються докази того, що порушення імунної регуляції грає роль у патогенезі обсесивно-компульсивного розладу (ОКР), синдрому Туретта та дитячих аутоімунних нейропсихіатричних розладів, пов'язаних зі стрептококовою інфекцією (PANDAS) .[34]
Історія
Здатність бачити та характеризувати різні нервові клітини, включаючи мікроглію, з'явилась у 1880 р., коли Франс Ніссл розробив варіант фарбування, який отримав назву «фарбування по Нісселю». Він і Ф. Робертсон вперше описали мікрогліальні клітини під час своїх гістологічних експериментів. Методи фарбування клітин у 1880 -х роках показали, що клітини мікроглії споріднені з макрофагами . Активацію мікроглії та утворення розгалужених мікрогліальних скупчень вперше відзначив Віктор Бабеш під час вивчення випадку сказу у 1897 році. Бабеш знайшов ці клітини при різних вірусних інфекціях мозку, але не знав, що він спостерігав кластери мікроглії.[35] Піо дель Ріо Хортега, учень Сантьяго Рамона-і-Кахаля, вперше назвав ці клітини «мікроглією» близько 1920 року. У 1927 р. він описав мікрогліальну реакцію на ураження мозку. У 1932 р. — описав «фонтани мікроглії», присутні в мозолистому тілі та інших перинатальних ділянках білої речовини. Після багатьох років досліджень Ріо Хортега став вважатися «батьком мікроглії».[36][37] Протягом тривалого періоду часу наші знання про мікроглію майже не збільшувались. Потім, у 1988 році, Хікі (Hickey) та Кімура (Kimura) показали, що периваскулярні мікрогліальні клітини походять з кісткового мозку і експресують високі рівні білків ГКГС ІІ класу, що використовуються для презентації антигену. Це підтвердило постулат Піо Дель Ріо-Хортеги про те, що мікрогліальні клітини функціонують подібно до макрофагів, здійснюючи фагоцитоз та презентацію антигену .
Примітки
- Ginhoux F, Lim S, Hoeffel G, Low D, Huber T (2013). Origin and differentiation of microglia. Frontiers in Cellular Neuroscience 7: 45. PMC 3627983. PMID 23616747. doi:10.3389/fncel.2013.00045.
- Lawson LJ, Perry VH, Gordon S (1992). Turnover of resident microglia in the normal adult mouse brain. Neuroscience 48 (2): 405–15. PMID 1603325. doi:10.1016/0306-4522(92)90500-2.
- Filiano AJ, Gadani SP, Kipnis J (August 2015). Interactions of innate and adaptive immunity in brain development and function. Brain Research 1617: 18–27. PMC 4320678. PMID 25110235. doi:10.1016/j.brainres.2014.07.050.
- Kreutzberg GW (March 1995). Microglia, the first line of defence in brain pathologies. Arzneimittel-Forschung 45 (3A): 357–60. PMID 7763326.
- Bushong EA, Martone ME, Jones YZ, Ellisman MH (January 2002). Protoplasmic astrocytes in CA1 stratum radiatum occupy separate anatomical domains. The Journal of Neuroscience 22 (1): 183–92. PMID 11756501. doi:10.1523/JNEUROSCI.22-01-00183.2002. Проігноровано невідомий параметр
|doi-access=(довідка) - Gehrmann J, Matsumoto Y, Kreutzberg GW (March 1995). Microglia: intrinsic immuneffector cell of the brain. Brain Research. Brain Research Reviews 20 (3): 269–87. PMID 7550361. doi:10.1016/0165-0173(94)00015-H.
- Dissing-Olesen L, Ladeby R, Nielsen HH, Toft-Hansen H, Dalmau I, Finsen B (October 2007). Axonal lesion-induced microglial proliferation and microglial cluster formation in the mouse. Neuroscience 149 (1): 112–22. PMID 17870248. doi:10.1016/j.neuroscience.2007.06.037.
- Kierdorf and Prinz, J Clin Invest. 2017;127(9):3201–3209. https://doi.org/10.1172/JCI90602.
- Cserép C, Pósfai B, Lénárt N, Fekete R, László ZI, Lele Z, Orsolits B, Molnár G, Heindl S, Schwarcz AD, Ujvári K, Környei Z, Tóth K, Szabadits E, Sperlágh B, Baranyi M, Csiba L, Hortobágyi T, Maglóczky Z, Martinecz B, Szabó G, Erdélyi F, Szipőcs R, Tamkun MM, Gesierich B, Duering M, Katona I, Liesz A, Tamás G, Dénes Á (January 2020). Microglia monitor and protect neuronal function through specialized somatic purinergic junctions. Science 367 (6477): 528–537. Bibcode:2020Sci...367..528C. PMID 31831638. doi:10.1126/science.aax6752.
- Hickman SE, Kingery ND, Ohsumi TK, Borowsky ML, Wang LC, Means TK, El Khoury J (2013). The microglial sensome revealed by direct RNA sequencing. Nature Neuroscience 16 (12): 1896–905. PMC 3840123. PMID 24162652. doi:10.1038/nn.3554.
- Block, M.L., Zecca, L. & Hong, J.S. Microglia-mediated neurotoxicity: uncovering the molecular mechanisms. Nat. Rev. Neurosci. 8, 57–69 (2007).
- Ji P, Schachtschneider KM, Schook LB, Walker FR, Johnson RW (2016). Peripheral viral infection induced microglial sensome genes and enhanced microglial cell activity in the hippocampus of neonatal piglets. Brain, Behavior, and Immunity 54: 243–251. PMC 4828316. PMID 26872419. doi:10.1016/j.bbi.2016.02.010.
- Horiuchi M, Smith L, Maezawa I, Jin LW (2017). CX3CR1 ablation ameliorates motor and respiratory dysfunctions and improves survival of a Rett syndrome mouse model. Brain, Behavior, and Immunity 60: 106–116. PMC 5531048. PMID 26883520. doi:10.1016/j.bbi.2016.02.014.
- Bergon A, Belzeaux R, Comte M, Pelletier F, Hervé M, Gardiner EJ, Beveridge NJ, Liu B, Carr V, Scott RJ, Kelly B, Cairns MJ, Kumarasinghe N, Schall U, Blin O, Boucraut J, Tooney PA, Fakra E, Ibrahim EC (2015). CX3CR1 is dysregulated in blood and brain from schizophrenia patients. Schizophrenia Research 168 (1–2): 434–43. PMID 26285829. doi:10.1016/j.schres.2015.08.010.
- Aloisi F (November 2001). Immune function of microglia. Glia 36 (2): 165–79. PMID 11596125. doi:10.1002/glia.1106.
- Christensen RN, Ha BK, Sun F, Bresnahan JC, Beattie MS (July 2006). Kainate induces rapid redistribution of the actin cytoskeleton in ameboid microglia. Journal of Neuroscience Research 84 (1): 170–81. PMID 16625662. doi:10.1002/jnr.20865.
- Davis EJ, Foster TD, Thomas WE (1994). Cellular forms and functions of brain microglia. Brain Research Bulletin 34 (1): 73–8. PMID 8193937. doi:10.1016/0361-9230(94)90189-9.
- Eggen BJ, Raj D, Hanisch UK, Boddeke HW (September 2013). Microglial phenotype and adaptation. Journal of Neuroimmune Pharmacology 8 (4): 807–23. PMID 23881706. doi:10.1007/s11481-013-9490-4.
- Lan X, Han X, Li Q, Yang QW, Wang J (July 2017). Modulators of microglial activation and polarization after intracerebral haemorrhage. Nat Rev Neurol 13 (7): 420–433. PMC 5575938. PMID 28524175. doi:10.1038/nrneurol.2017.69.
- Jelinek HF, Karperien A, Bossomaier T, Buchan A (1975). Differentiating grades of microglia activation with fractal analysis. Complexity International 12 (18): 1713–7. Архів оригіналу за 17 грудня 2008.
- Ferrer I, Bernet E, Soriano E, del Rio T, Fonseca M (1990). Naturally occurring cell death in the cerebral cortex of the rat and removal of dead cells by transitory phagocytes. Neuroscience 39 (2): 451–8. PMID 2087266. doi:10.1016/0306-4522(90)90281-8.
- Rissi, Daniel R.; Oliveira, Fabiano N.; Rech, Raquel R.; Pierezan, Felipe; Lemos, Ricardo A.A.; Barros, Claudio S.L. (2006). Epidemiologia, sinais clínicos e distribuição das lesões encefálicas em bovinos afetados por meningoencefalite por herpesvírus bovino-5 [Epidemiology, clinical signs and distribution of the encephalic lesions in cattle affected by meningoencephalitis caused by bovine herpesvirus-5]. Pesquisa Veterinária Brasileira (порт.) 26 (2): 123–32. doi:10.1590/S0100-736X2006000200010. Проігноровано невідомий параметр
|doi-access=(довідка) - Ritter MR, Banin E, Moreno SK, Aguilar E, Dorrell MI, Friedlander M (December 2006). Myeloid progenitors differentiate into microglia and promote vascular repair in a model of ischemic retinopathy. The Journal of Clinical Investigation 116 (12): 3266–76. PMC 1636693. PMID 17111048. doi:10.1172/JCI29683.
- Chung WS, Welsh CA, Barres BA, Stevens B (November 2015). Do glia drive synaptic and cognitive impairment in disease?. Nature Neuroscience (англ.) 18 (11): 1539–1545. PMC 4739631. PMID 26505565. doi:10.1038/nn.4142.
- Galloway, Dylan A.; Phillips, Alexandra E. M.; Owen, David R. J.; Moore, Craig S. (2019). Phagocytosis in the Brain: Homeostasis and Disease. Frontiers in Immunology (English) 0. ISSN 1664-3224. doi:10.3389/fimmu.2019.00790.
- Wolf A, Herb M, Schramm M, Langmann T (June 2020). The TSPO-NOX1 axis controls phagocyte-triggered pathological angiogenesis in the eye. Nature Communications 11 (2709): 2709. Bibcode:2020NatCo..11.2709W. PMC 7264151. PMID 32483169. doi:10.1038/s41467-020-16400-8. Проігноровано невідомий параметр
|doi-access=(довідка) - Banno M, Mizuno T, Kato H, et al (2005). The radical scavenger edaravone prevents oxidative neurotoxicity induced by peroxynitrite and activated microglia. Neuropharmacology (англ.) 48 (2): 283–90. PMID 15695167. doi:10.1016/j.neuropharm.2004.10.002.
- Ginhoux F, Prinz M (July 2015). Origin of microglia: current concepts and past controversies. Cold Spring Harbor Perspectives in Biology 7 (8): a020537. PMC 4526747. PMID 26134003. doi:10.1101/cshperspect.a020537.
- Zhu H, Wang Z, Yu J, Yang X, He F, Liu Z, Che F, Chen X, Ren H, Hong M, Wang J (March 2019). Role and mechanisms of cytokines in the secondary brain injury after intracerebral hemorrhage. Prog. Neurobiol. 178: 101610. PMID 30923023. doi:10.1016/j.pneurobio.2019.03.003.
- Gehrmann J (1996). Microglia: a sensor to threats in the nervous system?. Research in Virology 147 (2–3): 79–88. PMID 8901425. doi:10.1016/0923-2516(96)80220-2.
- Streit WJ (September 2006). Microglial senescence: does the brain's immune system have an expiration date?. Trends in Neurosciences 29 (9): 506–10. PMID 16859761. doi:10.1016/j.tins.2006.07.001.
- Mrak RE, Griffin WS (March 2005). Glia and their cytokines in progression of neurodegeneration. Neurobiology of Aging 26 (3): 349–54. PMID 15639313. doi:10.1016/j.neurobiolaging.2004.05.010.
- Pluvinage JV, Wyss-Coray T, et al. (11 квітня 2019). CD22 blockade restores homeostatic microglial phagocytosis in aging brains. Nature 568 (7751): 187–192. Bibcode:2019Natur.568..187P. PMC 6574119. PMID 30944478. doi:10.1038/s41586-019-1088-4.
- Frick, Luciana; Pittenger, Christopher (2016). Microglial Dysregulation in OCD, Tourette Syndrome, and PANDAS. Journal of Immunology Research 2016: 1–8. PMC 5174185. PMID 28053994. doi:10.1155/2016/8606057.
- Babeş, VM (1892). Certains caractères des lesions histologiques de la rage [Certain characteristics of the histological lesions of rabies]. Annales de l'Institut Pasteur (фр.) 6: 209–23.
- del Río Hortega, Pío; Penfield, Wilder (1892). Cerebral Cicatrix: the Reaction of Neuroglia and Microglia to Brain Wounds. Bulletin of the Johns Hopkins Hospital 41: 278–303.
- del Rio-Hortega F (1937). Microglia. Cytology and Cellular Pathology of the Nervous System: 481–534.