Жаба трав'яна
Жа́ба трав'яна́ (Rana temporaria) — земноводна тварина, один із представників роду бура жаба. Тварина середніх розмірів, досить типового для жаб вигляду, способу життя та поведінки. Розповсюджена практично в усій Європі (крім найпівденніших її районів), в Україні — переважно на заході, півночі та в центрі (в степових районах практично відсутня). Вид занесено до Додатку III «Конвенції з охорони дикої флори і фауни та природного середовища існування в Європі» (категорія «види, що підлягають охороні») та до «Червоного Списку хребетних Міжнародного союзу охорони природи (МСОП)».
? Трав'яна жаба | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 Трав'яна жаба | ||||||||||||||
| Біологічна класифікація | ||||||||||||||
| ||||||||||||||
| Rana temporaria Linnaeus, 1758 | ||||||||||||||
 | ||||||||||||||
Підвиди | ||||||||||||||
|
R.t.temporaria | ||||||||||||||
| Синоніми | ||||||||||||||
| Rana honnorati Héron-Royer, 1881 | ||||||||||||||
Посилання | ||||||||||||||
| ||||||||||||||

Морфологія

Жаби середніх розмірів (максимальна довжина тіла 100–110 мм). Загальне забарвлення тіла характеризується переважанням світло-коричневих тонів з наявністю плямистості. Як і для інших представників роду, доволі типовим є розвиток у потиличній області темної Λ-подібної (лямбдоподібної) плями та темних бічних «скроневих» смуг, що йдуть від кінчика морди майже до основи передніх кінцівок. На спинному боці тіла зазвичай виявлені темні плями неправильної форми, які в багатьох особин можуть збігатися з двома спинно-бічними складками шкіри. У деяких тварин уздовж середини спини проходить світла спинна смуга, яка зазвичай має нерізкі контури. На задніх кінцівках розвинені, як правило, поперечні смуги. Шкіра гладенька, з незначним розвитком дрібних горбочків.
Забарвлення черева представлене невеликими темними плямами неправильної форми на брудно-сірому або жовтуватому тлі, що торкаються одна одної. У шлюбний період горло самців набуває блакитного відтінку.
Порівняно з іншими бурими жабами задні кінцівки відносно короткі і якщо їх витягнути уздовж середньої осі тіла (у напрямку до голови), то скоковий (надступаковогомілковий) суглоб доходить лише до області очей або трохи заходить за них. Розміри (висота) внутрішнього п'яткового горбка невеликі, він виразно менший, наприклад, ніж в гостромордої жаби.
На внутрішньому боці перших пальців передніх лап самців розвинені шлюбні мозолі, розділені борозенками на чотири більш-менш виразні частки. Також наявні парні внутрішні горлові резонатори.
Поширення
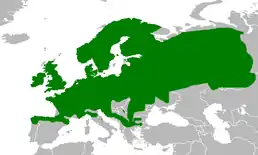
Вид поширений майже у всій Європі (за винятком центральної та південної частин Іберійського півострова, південної Італії та південних Балкан), на схід йде до Уралу, інтродукований в Ірландії та на Фарерських островах[1]. З 4-6 європейських підвидів трав'яної жаби в Україні живе номінативний — R.t.temporaria.
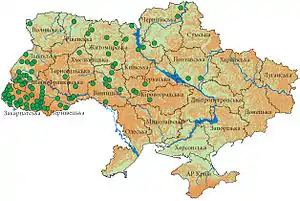
В Україні трав'яна жаба поширена у більшості північних, західних та східних регіонів, майже повністю відсутня у степових районах. На півдні межа поширення проходить північчю Одеської та Миколаївської (річка Чичиклія) областей, Кіровоградською областю (річка Громоклія), північчю Запорізької області, центральними районами Харківської та Дніпропетровської областей. На Лівобережжі R. temporaria майже повністю зникла, і зараз зрідка трапляється лише у Дніпровсько-Орільському заповіднику. Не виключено, що поширення цього виду може обмежуватися сильними весняними повенями, як, наприклад, її відсутність у заплаві річки Прип'ять[2].
У викопному стані трав'яна жаба виявлена у пізньоплейстоценових покладах у Тернопільській області, і в середньому пліоцені-середньому неоплейстоцені Одеської та Полтавської областей[3].
Мінливість
Матеріали з характеристики зовнішньоморфологічних ознак цих амфібій в межах української частини ареалу вкрай обмежені. Відомо, що закарпатські та прикарпатські вибірки з 12 ознак, що характеризують параметри тіла та кінцівок (7 абсолютних значень та 5 індексів), по 8 мають відміни, при цьому найбільші особини відзначені у більш високих ділянках оселення.
Розміри тіла трав'яних жаб у басейні Південного Бугу відразу ж після метаморфозу коливаються в межах 12,0-16,0 мм, а при відході на зимівлю 17-29 мм. Згодом довжина їхнього тіла на першому році життя складає в середньому 38,6-53,8 мм, на другому — 56,1-68,5 мм, на третьому — 68,8-77,2 мм, на четвертому — 77,3-84,5 мм, на п'ятому — 84,6-90,2 мм та на шостому — 90,8-94,4 мм[4].
Є відомості, що трав'яна жаба від інших видів більшою мірою відрізняється загальними розмірами тіла і меншою мірою — його пропорціями.
Судячи з результатів вивчення бурих жаб на території Росії (Нижній Новгород), підвищення антропогенного навантаження відбивається у зниженні розмірних характеристик (L., F., T., D. p. та ін.) і, крім того, збільшуються також відміни в розвитку морфологічних ознак на лівому та правому боках тіла тварин, тобто підвищується рівень флуктуаційної асиметрії.
При вивченні аномалій у тварин цього виду в Білорусі в 1977, 1978 і 1979 роках з'ясовано, що частота збільшення кількості пальців (полідактилія) в них становить 4,76, 3,14 і 2,92% відповідно.
Сучасних досліджень мінливості забарвлення та візерунку українських популяцій також бракує. Вивчення в інших регіонах частоти поширення різних типів візерунку на територіях з різним рівнем антропогенного тиску (Нижній Новгород, Росія) показало, що з підвищенням ступеню урбанізації зменшується кількість плямистих особин (морфа maculata), а чисельність крапчастих тварин (морфа punctata) зростає. Не виключено, що крапчастість та плямистість рецесивні щодо напівкрапчастості та напівплямистості.
В іншому регіоні (Піренеї, на висоті 2200 м) порівняння інтенсивності пігментації показало взаємозв'язок довжини тіла з кількістю чорних плям, що було більш чітко виявлено в самців.
Привертають до себе увагу дані, які свідчать про те, що плодючість цього виду у великих містах (Москва, Росія) більша, ніж у передмістях: в середньому 2 639 ікринок в одній кладці проти 1 700 відповідно[5]. Така сама залежність характеризує й розміри ікринок: 35,8 мкм проти 34,99 мкм на 17-й стадії розвитку (стадії розвитку виділяють залежно від ступеня розвитку починаючи від ікринки і до завершення метаморфозу[6]).
Генетика
Хромосомний набір трав'яної жаби (Україна, Івано-Франківська область, місто Яремче) представлений 26 хромосомами (2n = 26, NF = 52), серед яких виразно виділяються групи великих та дрібних хромосом. Більшість метафазних пластинок характеризуються наявністю вторинних перетяжок на довгому плечі 10-ї пари хромосом. Кількість та загальна морфологія хромосом не відрізняються від даних попередніх досліджень жаб цього виду на інших ділянках ареалу, наприклад, на території колишньої Югославії. При дослідженні каріотипу у жаб з цього регіону було показано наявність ядерцевих організаторів (NORs) на 10-й парі хромосом.
Порівняльно-генетичний аналіз трав'яної, прудкої та грецької жаб у Болгарії (Попов и др., 1996) дав підстави для висновку про більшу близькість R. temporaria до R. graeca та істотні відміни від R. dalmatina. В іншому схожому порівнянні трав'яної жаби з сімома 26-хромосомними видами цієї групи (R. japonica, R. tsushimensis, R. okinavana, R. longictus, R. asiatica, R. amurensis, R. latouchii) було показано, що величина генетичної відстані між ними коливається в межах 0,294-2,913[7].
Біотопи та чисельність

Вид приурочений до лісових та лісостепових ділянок, але на відміну від зелених жаб з водоймами пов'язаний тільки в період розмноження. Дані про чисельність у різні сезони активності і в різних біотопах відрізняються. Так, у Закарпатті чисельність жаб у березні на нерестовищі в одній водоймі площею 2,5 м² досягала 37 особин, в іншому — 100 особин на 100 м берегової лінії[8]. За результатами обліку після закінчення сезону розмноження (червень-вересень), поширення трав'яних жаб на 100 м маршруту становила 3-20 особин.
У басейні річки Південний Буг у сезон розмноження найвища щільність трав'яних жаб відзначається у ставках з прилеглими лісовими біотопами: до 103–116 особин на гектар водойми. На нерестовищах, розташованих в антропогенних ділянках (поля), чисельність становить 77-83 особин на гектар.
Дані обліків у різних біотопах і у різні періоди річної активності відрізняються і непрямо вказують на існування міграцій. Так, у названому вище регіоні (басейн річки Південний Буг) влітку в широколистяних лісах щільність трав'яних жаб становить 32-40 особин/га, а восени вона тут досягає величини 109–122 особин/га, на зволожених луках щільність з 84-94 особин/га спадає до 7-9 особин/га.
На інших ділянках ареалу (Карелія та Архангельська область Росії) цей показник коливається від 0,19 до 10,33 та від 0 до 8,80 особин/га відповідно.
Особливості біології
Для трав'яних жаб характерна присмерково-нічна активність. Удень вони ховаються у порожнинах під камінням, пеньками, у листовій підстилці. При активному полюванні та температурі приземного шару повітря в 11-21°С, температура тіла тварини на 1-2,5°С вища.
Поява трав'яних жаб у весняний період залежить від погодних особливостей року, та все ж вона припадає на один з найбільш ранніх строків у порівнянні з іншими земноводними. Так, на Закарпатті (околиці Ужгорода), вони активні вже на початку — середині березня, коли у водоймах ще є залишки криги. Із збільшенням висоти місцевості над рівнем моря весняна поява зсувається на пізніші строки.
Живлення
Доросла трав'яна жаба харчується різноманітними безхребетними відповідного розміру[9]. Улюбленою їжею є комахи (особливо мухи), равлики, слимаки і черви). Жаби полюють з допомогою довгого та липкого язика. Їхній раціон доволі сильно змінюється протягом життя. Дорослі особини харчуються лише на суходолі, ювенальні особини можуть також харчуватись у воді. Пуголовки переважно травоїдні. Вони харчуються водоростями, детритом і деякими вищими рослинами, але також можуть полювати і на невеликих тварин[10]. Трав'яна жаба не харчується протягом шлюбного сезону.
Розмноження

Парування після весняного прокидання починається в середньому через 2-6 діб. У цей час самці можуть намагатися охопити будь-який предмет, що рухається в межах їхнього зору, через що виникає амплексус (схоплення особини самцем для спаровування) з особинами інших видів безхвостих амфібій. Тривалість нересту складає в середньому всього 3-7 діб, при цьому самці залишаються у водоймах на декілька днів довше від самиць.
Вибір придатних водойм для нересту зумовлений ландшафтними особливостями, та найчастіше це слабко проточні або стоячі водойми у біотопах з деревною або деревно-чагарниковою рослинністю. R. temporaria для відкладання ікри може використовувати також водойми у трансформованих біотопах з прилеглими полями.
Дослідження гідрохімічних показників на нерестовищах у Львівській області показало, що їм притаманна нейтрально-лужна реакція води (рН до 7-7,6), вміст кисню 2-6 мг/л, загальна мінералізація 320–480 мг/л.

Трав'яні жаби, як і чимало інших видів безхвостих амфібій, характеризуються груповою кладкою, що виявляється у зосередженні на місцях розмноження багатьох десятків пар тварин. Звичайно пік розмноження трав'яних жаб настає на декілька днів раніше, ніж у R. arvalis, що живуть у тому ж регіоні.
Існує залежність між кількістю ікринок та віком та розмірами самиці: трирічні самиці при довжині тіла близько 62 мм відкладають приблизно 930 ікринок, а шестирічні при розмірах тулуба 87 мм — понад 3660 ікринок. Така сама залежність встановлена й для розмірів ікринок — від самиць трирічного віку вони мають середній діаметр близько 1,5 мм, шестирічного — близько 2,5 мм.
Розмір кладки на Закарпатті до 4 000 ікринок, в інших ділянках ареалу (басейн річки Кама, Росія) дещо нижчий і складає в середньому близько 1 750. Ще далі на північ (Соловецькі острови, Біле море, Росія) середня кількість ікринок у кладці знижується до 506. Ікра доволі холодостійка й зберігає життєздатність навіть з появою криги при весняних заморозках. Ікринки після набрякання оболонок спливають, звичайно утворюючи великі скупчення. Період ембріонального розвитку чимало залежить від температури середовища, але найчастіше триває 7-9 днів, а загальний час метаморфозів за лабораторних умов при температурі води 17-19°С становить 49-55 днів. В умовах долини річки Південний Буг середня тривалість личинкового розвитку становить приблизно 58 днів.
Розвиток ікри та пуголовків


Тривалість личинкового розвитку в центральних та південних регіонах Росії відрізняється — метаморфози на півдні тривають 85-90, а на півночі 21-60 діб.
Температура всередині агломерацій ікри (скупченнях декількох кладок в одному місці) на 1-4°С вища, ніж у навколишньому шарі. При цьому загибель ікри до вилуплення личинок у зовнішніх частинах таких агломерацій становить 10%, а всередині — до 50%. Таким чином, специфікою таких кладок є як захист від різких падінь температури, так і одночасне зростання загибелі внаслідок погіршення аерації та накопичення продуктів життєдіяльності. При цьому зміщення показника рН у бік підвищення кислотності призводять до більшої загибелі зародків. Разом з тим частина потомства гине незалежно від характеристик водойми завдяки своїй ослабленості.
Для трав'яних жаб, як і для багатьох інших видів безхвостих земноводних, характерний високий рівень загибелі на ранніх етапах розвитку, і зазвичай з усієї кількості відкладених ікринок (за матеріалами дослідження на території колишнього СРСР) виживає не більше 4% цьоголітків). У трьох штучно створених ставках у Німеччині кількість молоді, що виживала, коливалася від 0,2 до 59,2 цьоголітків на 1 самицю, а в перерахунку на ікру — від 0,00017 до 0,045 цьоголітків на 1 самицю.
Чисельність трав'яних жаб, що беруть участь у розмноженні, становить приблизно 1% початкової генерації. При цьому на ембріональній стадії гине 30-50%, у період вилуплення, коли пуголовки перебувають ще в ікряних оболонках, відбувається загибель ще 30% генерації, і далі аж до метаморфозів чисельність знижується на 83-97% загальної кількості відкладених ікринок. Це свідчить про те, що основним фактором регуляції чисельності є «внутрішньопопуляційні взаємодії», а не хижаки чи їжа.
Якщо личинки трав'яної жаби у водоймах є фізіологічно дещо старшими, то вони пригнічують (інгібують) ріст і розвиток личинок R. arvalis і Р. esculentus. При підвищеній щільності це може призвести до повного вимирання пуголовків двох останніх видів.
Розміри личинок відразу після виходу з ікринок становлять 6-8 мм, перед метаморфозом — до 46 мм (з хвостом). Вилуплені личинки чорного кольору, довжина хвоста перед метаморфозом приблизно удвічі перевищує довжину тулуба. В умовах центральної України довжина тіла личинок перед метаморфозом дещо більше — 42 мм.
У віковій структурі українських популяцій цьоголітки становлять 47,3%, чисельність жаб різних вікових груп коливається: 1-ї вікової групи — 26,4%, 2-ї — 11,1%, 3-ї — 9,7%, 4-ї — 2,8% та 5-ї — близько 2% (в основу виділення вікових груп покладено дані з мічення та вивчення мінливості довжини тіла).
Статева зрілість та нерестові міграції
Трав'яні жаби стають статевозрілими у віці 2-3 роки при довжині тіла близько 50 мм. Разом з тим у Польщі (околиці Кракова), серед 402 самиць, спійманих на зимівлі, було виявлено дві особини довжиною 6 і 7 см, які за таких розмірів виявилися не статевозрілими. Довжина тіла статевозрілих самиць коливається в межах 5,5-10,5 см (в 72,8% від 7 до 8,5 см), самців 5,4-9,7 см (в 84,2% від 6,4 до 8,4 см).
У білоруських популяціях (Мінська область) мінімальний вік статевозрілих особин як самців, так і самиць, — 3 роки, максимальний — 7 років. При просуванні на північ (околиці Санкт-Петербурга, Росія) максимальна тривалість життя зменшується до 6 років, а чисельність трирічок у популяції становить 47% статевозрілих. У східній частині ареалу (Середній Урал) чисельність у популяції трирічних особин спадає до 18%, але максимальна тривалість життя збільшується до 8 років. Ці ж показники на Полярному Уралі дорівнюють 70% і 17 рокам відповідно.
На особливості біології цього виду (початок відкладання ікри, строки вилуплення пуголовків, метаморфоз тощо) істотний вплив справляє не тільки температура, але й, що особливо важливо, попередні фенологічні явища: такі як температура лютого та березня, строки танення снігу, червневі коливання температури тощо.
Матеріали з мічення та повторних відловів трав'яних жаб в околицях Львова свідчать про існування тривалих міграцій від місць розмноження на місця постійного пробування в літній період.
У Болгарії також було вказано на існування нерестових міграцій униз течією річки (катадромна міграція), а після відкладення ікри — проти течії (анадромна міграція). Катадромні міграції відбуваються навесні разом з паводковими водами на відстань 8-10,5 км за 3-5 днів. Анадромні міграції тривають набагато довше, і на вихідні місця жаби повертаються лише у середині літа (початок нересту в середині січня — кінці лютого). Причина цих міграцій пов'язана з прив'язаністю R. temporaria до вихідної водойми. Так, вюрмське потепління відтіснило їх у гори, а брак придатних для нересту місць став причиною щорічних нерестових міграцій.
З іншого боку, більша частина (61%) повторно мічених жаб у північно-східній Чехії була піймана в межах 20 м від того місця, де їх випускали.
Зимівля
Сезон активності завершується у жовтні-листопаді на рівнинах і в жовтні у горах. Зимівля не статевозрілих особин може відбуватися на суходолі в норах гризунів, старих пнях, купах листя тощо. Дорослі особини зимують зазвичай у незамерзлих водоймах при рН води 5,6-5,8.
У зимових агрегаціях R. temporaria може утворювати три різні типи скупчень. Перший («щільний») характеризує скупчення не статевозрілих жаб з довжиною тіла 21,8-48,8 мм у незамерзлих струмках з температурою води 2,4-5,2°С. Другий тип («розсіяний») властивий статевозрілим особинам у періодично замерзаючих струмках (температура 0,3-1,7°С), де жаби розосереджені на ділянці водойми завдовжки близько 20 м. Третій тип («проміжний») є комбінованим і характеризується зимівлею у періодично замерзаючих джерельних ямах з температурою води 2,2-4,2°С та наявністю як статевозрілих, так і не статевозрілих особин. Існують дані про те, що скупчення трав'яних жаб у водоймах на зимівлі (температура води тут близько 4 °C) можуть досягати декількох тисяч особин, а самі такі скупчення інколи мігрують на 100–120 м.
Природні вороги
Серед прямих ворогів на стадії ікри та личинки слід вказати на звичайного та гребінчастого тритонів, червоночереву джерелянку, озерних та ставкових жаб. У Підмосков'ї (Росія) одним з факторів, дія якого з початку 1990-х років стала знижувати тут чисельність трав'яних жаб, є карась, а саме те, що ним були заселені місцеві водойми. Так, в умовах лабораторного експерименту менше десяти особин середніх розмірів повністю виїдали личинок R. temporaria протягом 2-3 діб після вилуплення. Після проходження метаморфозу їх поїдають також і інші риби (ротань-головешка, окунь, щука, сом та інші), звичайна гадюка і, особливо часто, звичайний вуж, багато видів птахів (крук, сіра ворона, сорока, сорокопуди тощо) та ссавці (ласка, норка, лисиця, вовк та інші). Є дані про те, що основними ворогами цьоголітків є землерийки (зокрема, білозубки й мідиця).
Примітки
- Rana temporaria have established themselves as a wild population in Nólsoy
- Боркин Л. Я., 1998. Класс Амфибии, или Земноводные. — В кн. Ананьева Н. Б., Боркин Л. Я., Даревский И. С., Орлов Н. Л. Земноводные и пресмыкающиеся. Энциклопедия России. — М. : ABF.— C.19-174.
- Ратников В. Ю., 2002. Позднекайнозойские земноводные и чешуйчатые пресмыкающиеся Восточно-европейской равнины. — Тр. Научно-иссл. Ин-та геологии Воронеж. гос. ун-та. — 10. — С. 1-138.
- Гончаренко Г. Є., 2002. Земноводні Побужжя. Монографія. — Київ : Наук. світ.— 219 с.
- Северцова Е. А., 2001. Плодовитость отромордой (Rana temporaria) и травяной (R. arvalis) лягушек в Москве и Подмосковье. — В кн.: Вопросы герпетологии. Материалы 1-го съезда Герпетологического общества им. А. М. Никольского (Пущино-на-Оке, 4-7 декабря 2000 г.) — Пущино; М. — С. 257–259.
- Дабагян Н. В., Слепцова Л. А., 1975. Травяная лягушка Rana temporaria L. В кн.: Объекты биологии развития. — М. : Наука. — C. 442–462.
- Nisihioka M., Sumida M., Borkin L. J., Wu Z., 1992. Genetic differentiation of 30 populations of 12 brown frog species distributed in the Palearctic region elucidated by the electrophoretic method. — Sci. Rep. Lab. Amphibian Biol., Hiroshima Univ.— 11: 109–160.
- Щербак Н. Н., Щербань М. И., 1980. Земноводные и пресмыкающиеся Украинских Карпат. — Киев : Наук. думка. — 268 с.
- Трав'яна жаба на вебсайті BBC. Додано 2007-08-09.
- Sergius L., Kuzmin (1999-11-10). «Rana temporia». AmphibiaWeb. Retrieved 2007-08-09.
Джерела та література
- Писанець Є. Земноводні України (посібник для визначення амфібій України та суміжних країн). — Київ : Вид-во Раєвського, 2007. — 197 с.
- Гончаренко Г. Є. Земноводні Побужжя. — Київ: Наук. світ, 2002.— 219 с.
- Боркин Л. Я. Класс Амфибии, или Земноводные // Земноводные и пресмыкающиеся. Энциклопедия России. — Москва: ABF, 1998.— C.19-174.
- Северцова Е. А. Плодовитость отромордой (Rana temporaria) и травяной (R. arvalis) лягушек в Москве и Подмосковье // Вопросы герпетологии. — Пущино; Москва, 2001. — С. 257–259.
Посилання
- Сторінка виду на сайті Червоного Списку МСОП
- Карта розповсюдження виду за даними МСОП
- Сторінка виду на сайті Amphibiaweb
|
|
Ця стаття належить до вибраних статей Української Вікіпедії. |