Ранні міграції людства
Ранні міграції людства - найбільш ранні міграції та розширення ареалу архаїчної та сучасної людини по різних континентах, які почалися 2 мільйони років тому міграцією "з Африки" Homo erectus. Після цієї початкової міграції відбулися міграції інших архаїчних людей, включно з H. heidelbergensis, які жили бл. 500 тис. років тому та ймовірно були предками і денисівської людини, і неандертальців. Вважається, що ранні гомініди "перетинали суходольні мости, які пізніше стали вкриті водою" (History Alive, надр. 2004, TCI).
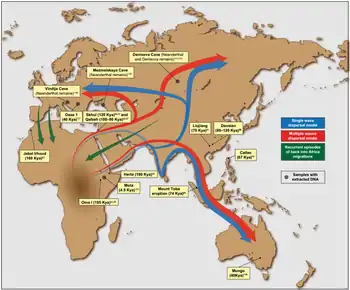
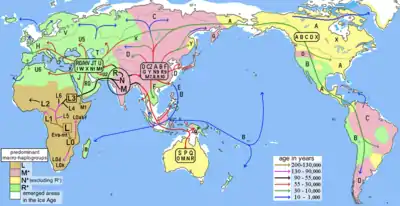
Homo sapiens розселилися Африкою невдовзі після виділення в окремий вид, бл. 300 тис. років тому.[1] "Теорія африканського походження людини" припускає, що анатомічно сучасні люди за межами Африки є нащадками популяції Homo sapiens, які мігрувала зі Східної Африки десь 70 тис. років тому та поширилася вздовж південного берега Азії до Океанії раніше за 50 тис. років тому. Сучасні люди поширились Європою бл. 40 тис. років тому.
Відомо, що мігруючі популяції сучасних людей змішувались з місцевими різновидами архаїчних людей, тому сучасні нам людські групи незначною мірою є нащадками (менше 10% внеску) регіональних підвидів архаїчних людей.[2]
Після останнього льодовикового максимуму, північно-євразійські популяції мігрували до Америк бл. 20 тис. років тому. Північна Євразія була заселена не раніше 12 тис. років тому, на початку Голоцену. Арктична Канада та Гренландія були заселені палеоескімосами бл. 4 тис. років тому. І нарешті Полінезія була заселена 2 тис. років тому, експансією австронезійців.
Ранні люди (до Homo sapiens)
Найраніші люди розвинулись з предків-австралопітекоподібних десь 3 млн. років тому, найбільш ймовірно у Східній Африці, довкола Кенійської рифтової долини, де були знайдені найстаріші відомі кам'яні знаряддя. Кам'яні знаряддя, нещодавно відкриті на ділянці Шангчжен (Shangchen) у Китаї та датовані 2,12 млн.років тому, вважаються найбільш раннім відомими свідченням гомінідів за межами Африки, випереджаючи Дманісі у Грузії на 300 тис. років.[3]
Homo erectus
У періоді між 3 і 2 млн.років тому, Homo поширились по Східній Африці та до Південної Африки (Telanthropus capensis), але не до Західної Африки.
Бл. 1,9 млн.років тому, Homo erectus мігрувала з Африки через Левантійський коридор і Сомалійський півострів до Євразії. Цю міграцію пов'язують з роботою Сахарського насосу, бл. 1,9 млн.років тому.
Homo erectus поширився по більшій частині Старого світу, досягнувши аж до Південносхідної Азії. Це поширення прослідковується за олдувайською кам'яною індустрією, яка 1,3 млн. років тому простягалась на північ аж до 40-ї паралелі (Сяочанліянг (спрощ.: 小长梁; кит. трад.: 小長梁; піньїнь: Xiǎochángliáng)).
Ключовими ділянками свідчення цієї ранньої міграції з Африки є Ріват у Пакистані (~2 млн.років тому?[4]), Убейдія у Леванті (1,5 млн.років тому) і Дманісі на Кавказі (1,81 ± 0,03 млн. років тому, p = 0.05[5]).
Китай був заселений десь 1,66 млн. років тому, на підставі кам'яних артефактів знайдених у долині Ніхеван (Nihewan).[6] Археологічна ділянка Сіходу (西侯渡) у провінції Шаньсі містить найбільш ранні відомі свідчення освоєння вогню Homo erectus, що датуються 1,27 млн. років тому.[7]
Південно-Східну Азію (острів Ява) ранні люди досягли бл. 1,7 млн. років тому (Мегантроп). Західну Європу було вперше заселено бл. 1,2 млн. років тому (Атапуерка).[8]
Роберт Беднарік запропонував контроверсійну теорію, що Homo erectus могли будувати плоти та перетинати океани.[9]
Після H. erectus
Через мільйон років після виокремлення, H. erectus почав розподіл на нові види. H. erectus є хроновидом і ніколи не вимирав, тому його "пізнє виживання" є предметом таксономічного звичаю. Вважається, що пізні форми H. erectus існували до близько 500 тис. років тому або до 143 тис.років тому найпізніше,[10] з похідними формами, такими як H. antecessor у Європі бл.800 тис. років тому та H. heidelbergensis у Африці бл. 600 тис. років тому. Остання в свою чергу поширилась східною Африкою (H. rhodesiensis) та до Євразії, де стала предком неандерталькців і денисівської людини.
H. heidelbergensis, неандертальці та денисівці поширились на північ вище 50-ї паралелі (Ерсхем Піт, Боксгроув бл.500 тис. років тому, Свонскомб Херітедж Парк 400 тис. років тому, Денисова печера 50 тис. років тому). Припускається, що неандертальці могли досягнути межі Арктики бл. 32 тис. років тому, коли їх витіснили з їх більш ранніх територій H. sapiens, на підставі розкопок 2011 року на ділянці Бузовая в Уральських горах (Республіка Комі, 65.02° пн. ш. 57.42° сх. д.).[11]
Вважається, що інші архаїчні види людей поширились в цей час Африкою, хоча скам'янілостей знайдено мало. Їх присутність припускається на основі ознак схрещування з сучасними людьми, які знаходять у геномі африканських народів.[12][13][14][15] Homo naledi, знайдений у Південній Африці 2013 року та попередньо датований бл. 300 тис. років тому, може бути скам'янілістю-доказом існування таких архаїчних видів людей.[16]
Неандертальці поширились на Близький Схід та Європу, а той час як денисовці ймовірно поширились Центральною та Східною Азією аж до Південно-Східної Азії та Океанії. Існують свідчення, що денисовці схрещувались з неандертальцями у Центральній Азії, де перетинались їх ареали.[17]
Найбільш ймовірним вважається, що H. sapiens розвинулась з африканського підвиду H. antecessor бл. 300 тис. років тому.[18]
Homo sapiens
Поширення Африкою
Вважається, що Homo sapiens (анатомічно сучасна людина) виникла бл. 300 тис. років тому, на основі термолюмінісцентного датування артефактів з Джебель-Ірхуд, Марокко, результати якого були надруковані 2017 року.[19] До цього, найстарішими відомими залишками анатомічно сучасної людини тривалий час вважалися залишки Омо, розкопані між 1967 і 1974 роками у Національному парку Омо, Ефіопія, і датовані 200 тис. років тому.[20]
Ранні сучасні люди поширились до Західної Євразії, Центральної, Західної та Південної Африки з часу їх виділення у вид. Хоча таке раннє поширення у Євразію, ймовірно, не було успішним,[21][17] поширення до Південної і Центральної Африки мали наслідком найбільш ранніх за часом відмінностей у сучасній людські популяції. Раннє поширення сучасної людини в Субсахарській Африці, ймовірно, спричинили припинення пізньої Ашельської культури (Культура Фаурсміт) бл. 130 тис. років тому, хоча деякі дослідники припускають дуже пізнє співіснування архаїчної та сучасної людини (аж до 12 тис. років тому), зокрема у Західній Африці.[22]
Предки сучасних койсанських народів поширилися до Південної Африки не пізніше 150 тис. років тому, можливо навіть раніше 260 тис. років тому,[23] і на початок так званої MIS 5 "мегапосухи" 130 тис. років тому існувало два кластери популяції предків у Африці, які були носіями мтДНК гаплогрупи L0 у південній Африці, що є предками койсанських народів, і носіями гаплогрупи L1-6 у центральній/східній Африці, які є предками решти людства. Існувала значна зворотня міграція носіїв гаплогрупи L0 у східну Африку між 120 і 75 тис. років тому.[24]
Поширення у центральну Африку предків центрально-африканських популяцій збирачів (африканських пігмеїв) найбільш ймовірно відбулося раніше 130 тис. років тому, і точно раніше 60 тис. років тому.[25]
Ситуація у Західній Африці важко піддається інтерпретації з огляду на малу кількість виявлених скам'янілостей. Ймовірно Homo sapiens досягнули західну Сахельську зону до 130 тис. років тому, а тропічні Західно-Африканські археологічні ділянки, які пов'язують з H. sapiens, з'явились лише після 130 тис. років тому. На відміну від решти Африки, архаїчні ділянки Середньої Кам'яної доби існували тут дуже довго, аж до межі Голоцену (12 тис. років тому), вказуючи на можливість виживання археантропів та їх пізню гібридизацію з H. sapiens у Західній Африці.[22]
Раннє поширення Північною Африкою
Популяції H. sapiens мігрували до Леванту та Європи між 130 і 115 тисяч років тому, а можливо і у більш ранніх хвилях, аж до 185 тис. років.[21] Вважається, що ці ранні міграції не призвели до тривалої колонізації та відкотились бл. 80 тис. років тому.[17] Існує ймовірність, що ця перша хвиля експансії досягла Китаю (або навіть Північної Америки[26]) не пізніше 125 тис. років тому, але вимерла, не залишивши сліду у геномі сучасних нам людей.[17]
Також існують певні докази на користь того, що сучасні люди полишили Африку не пізніше 125 тис. років тому двома різними шляхами: через долину Ніла на Середній Схід, принаймні до сучасного Ізраїлю (Кафзех: 120–100 тис. років тому); та другий шлях через сучасну Баб-ель-Мандебську протоку Червоного моря (яка в той час була набагато мілкіша та вужча), перетнувши її до Аравійського півострова і розселившись по території сучасних ОАЕ (125 тис. років тому)[27] and Oman (106,000 years ago)[28], а потім, ймовірно, далі мігрували на Індійський субконтинент (Джавалпурам: 75 тис. років тому). Незважаючи на те, що в цих трьох місцях ще не знайдені людські залишки, видима схожість між кам'яними знаряддями, знайденими у Джебел Файя,, зі знаряддями з Джавалпурама та деякими африканськими дозволяє припустити, що їх творці всі були сучасними людьми .[29] Ці знахідки можуть підтримати твердження, що сучасна людина прибула з Африки у південний Китай бл. 100 тис. років тому (печера Чжижень, Чжиженьдун, місто Чунцзо: 100 тис. років тому;[30] та люцзянський гомінід (провінція Люцзян): контроверсійно датований 139–111 тис. років тому[31]). Результати датування зубів з Лунадонг (долина Бубінг, Гуансі-Чжуанський автономний район, південний Китай), які включають правий верхній другий корінний зуб та лівий нижній другий корінний зуб, вказують на те, що ці зуби можуть мати вік 126 тис. років.[32][33]
Оскільки ці попередні виходи з Африки не залишили слідів у результатах генетичного аналізу на основі Y-хромосоми та на основі мтДНК (які представляють лише незначну частину людського генетичного матеріалу), ймовірно ті сучасні люди не вижили або вижили лише у незначній кількості та були асимільовані нашими основними предками. Поясненням їх вимирання (або малого генетичного сліду) може бути теорія катастрофи вулкана Тоба (74 тис. років тому). Однак, деякі дослідники доводять, що вплив цієї катастрофи на людську популяцію не був драматичним.[34]
Азіатський центр походження та поширення мтДНК-гаплогрупи L3 також припускається на основі виявлених скам'янілостей, схожих дати формування L3 та її похідних клад M і N, поширених Євразією (~71 тис. років тому), віддаленого розташування у Південно-Східній Азії найстаріших субклад M і N, та порівняного віку батьківської гаплогрупи DE. Таким чином пропонується, що після початкової міграції з Африки ранньої анатомічно сучасної людини бл. 125 тис. років тому, повністю сучасні жінки - носії L3-гаплогрупи мігрували назад до місця виникнення материнської гаплогрупи у Євразії бл. 70 тис. років тому разом з чоловіками - носіями батьківської гаплогрупи E, виникнення якої також припускається в Євразії. Також припускається, що ці нові евразійські роди значною мірою замінили старі автохтонні чоловічі та жіночі африканські роди.[35]
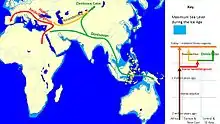
Прибережна міграція
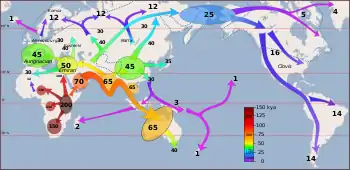
Так зване "нещодавнє поширення" сучасних людей розпочалося бл.70 тис.років тому; саме воно призвело до триваючого розселення сучасних людей по всьому світу.
Невелика група людей (що ймовірно нараховувала менше 1000 людей), що були носіями мітохондріальної гаплогрупи L3, з популяції, яка населяла Східну Африку,[36][37] перетнула Червоне море через Баб-ель-Мандеб, і досягла сучасного Ємену, десь після 75 тис. років тому.[38] Нещодавній перегляд історичних доказів також надав підтримку існуванню більш північного шляху через Синай/Ізраїль/Сирію (Левант).[17] Їх нащадки поширились вздовж прибережного маршруту довкола Аравії та Персії до Індійського субконтиненту не пізніше 55 тис. років тому. Сценарій прибережної міграції у періоді між бл. 70 і 50 тисячами років тому асоціюється з появою мітохондріальних гаплогруп M і N, які обидві є похідними від L3.
Фрагмент щелепи з вісьмома зубами, знайдений у печері Міслія, Ізраїль, був датований бл.185 тис. років тому. Шари ґрунту у цій печері, які були датовані між 250 і 140 тис. років тому, містили знаряддя левалуазького типу, що може змістити датування першої міграції на ще більш ранній період, якщо ці знаряддя будуть пов'язані зі знахідками щелеп сучасної людини.[39][40]
На своєму шляху H. sapiens змішувалися з неандертальцями та денисівською людиною,[41] свідченням чого є те, що ДНК денисівської людини складає 0,2% ДНК мешканців суходольної Азії та корінних американців.[42]
Океанія
Міграції продовжувались вздовж узбережжя Азії до Південно-Східної Азії і Океанії, і Австралія була колонізована не пізніше 50 тис. років тому.[43] Досягнувши Австралії, H. sapiens вперше поширився далі, ніж H. erectus.
Спадок денисівської людини прослідковується у меланезійців, австралійських аборигенів та у менших розкиданих груп народів Південно-Східної Ахії, таких як Маманва, негритоського народу на Філіппінах, що дозволяє припустити схрещення у Східній Азії, де жила Денисівська людина.[44][45][46] Денисівська людина могла перетнути лінію Воллеса, і Воллесія могла стати її останнім рефугіумом.[47][48] Homo erectus перетнув протоки Ломбок, досягнувши острова Флорес, але не досягли Австралії.[49]
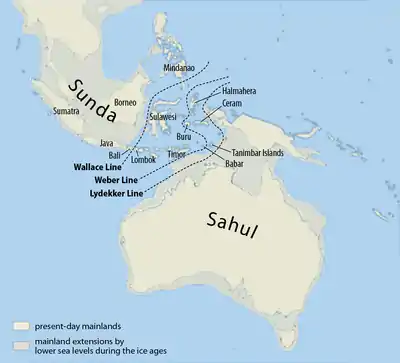
Коли рівень моря був значно нижчим, більшість морської Південно-Східної Азії утворювало одну масу суходолу, відому як Зонда. Міграція продовжувалась на південний схід прибережним маршрутом до проток між Зонда та Сахул (континентального суходолу, який включав сучасні Австралію та Нову Гвінею. Діри у лінії Вебера становлять до 90 км у ширину,[50] тому міграція до Австралії та Нової Гвінеї потребувала б навичок плавання морем. Міграція також продовжувалась вздовж берега, повернувши врешті-решт на північний схід до Китаю та досягнувши Японії, після чого вона повернула всередину континенту. Свідченням цього є патерн мітохондріальних гаплогруп, які походять від гаплогрупи M, та у Y-хромосомній гаплогрупі C.
Розшифровка генома аборигена зі старого зразка волосся у Західній Австралії виявила, що ця особа була нащадком людей, які мігрували до Східної Азії між 75 і 62 тис. років тому. Це підтримує теорію одиничної міграції до Австралії та Нової Гвінеї до прибуття сучасних азійців (між 38 і 25 тис. років тому) та їх пізнішої міграції до Північної Америки.[51] Вважається, що ця міграція сталась бл. 50 тис. років тому, до того, як Австралія та Нова Гвінея були відділені зростанням рівня моря бл. 8 тис. років.[52][53] Це підтримується датуванням свідчень найстарішого відомого поселення в Австралії 50–60 тис. років тому,[43][54] найстаріших відомих залишків людини - 40 тис. років тому,[43] найстарішими відомими залишками людини, датованими принаймні 65 тис. років тому[55] та вимиранням Австралійської мегафауни між 46 і 15 тис. років тому, яке на думку Тіма Фланнері сталося внаслідок діяльності людини[56] аналогічно до того, що відбулося в Америках. Предметом дискусії залишається, чи продовжувалось в Австралії використання знарядь Кам'яної доби.[57]
Поширення Євразією
Вважається, що популяція, яка досягла Південної Азії внаслідок прибережної міграції, лишилась там орієнтовно на 10 тисяч років - між 60 і 50 тис. років тому, перш ніж поширитись далі Євразією. Поширення ранніх людей на початку Пізнього палеоліту, спричинило появу великих популяційних груп Старого світу і Америк.
На заході популяції Пізнього палеоліту асоціюються з мітохондріальною гаплогрупою R та її похідними, які поширені в Азії та Європі, зі зворотньою міграцією гаплогрупи M1 до Північної Африки та Сомалійського півострову декілька тисячоліть тому.
Датування присутності людини у Європі вважається точним після 40 тис. років тому, та ймовірно 43 тис. років тому,[58] коли вона швидко замінила там популяцію неандертальців. Сучасні європейці мають спадок неандертальців, але вважається, що значне змішування між неандертальцями та сучасною людиною завершилось не пізніше 47 тис. років тому, тобто відбувалось до прибуття сучасної людини в Європу.[59]
На основі дослідження мітохондріального ДНК вважається, що сучасна людина пройшла принаймні через одне генетичне пляшкове горло, коли була значно зменшена різноманітність геному. Генрі Гарпендінг (Henry Harpending) припустив, що люди поширились з географічно обмеженої території бл. 100 тис. років тому, пройшли через генетичне пляшкове горло, а потім бл. 50 тис. років тому розпочалось різке зростання географічно розкиданих популяцій, починаючи з Африки і поширившись і інші місця.[60] Існують кліматологічні та геологічні свідчення на користь пляшкового горла. Вибух вулкану Тоба створив холодний період тривалістю 1000 років (це було найбільшим вулканічним виверженням Четвертинного періоду, можливо зменшивши людські популяції до кількох тропічних рефугіумів. Припускається, що загальна популяція людини зменшувалась до 15 тисяч. У таких умовах генетичний дрифт та ефект засновника могли максимізуватись. Більша різноманітність серед африканських геномів може частково завдячувати більшій кількості африканських рефугіумів під час тривання катастрофи Тоба.[61]
Однак, нещодавній огляд підкреслює, що гіпотеза про один виток неафриканських популяцій менше узгоджується з аналізом стародавнього ДНК, ніж гіпотеза про багато джерел з генетичним змішуванням по Євразії.[17]
Європа
Вважається, що анатомічно сучасна людина досягла Європи бл.40 тис. років тому з Центральної Азії та Середнього Сходу в результаті адаптації до полювання на великого звіра фауни післяльодовикового степу.[62] Неандертальці були присутні на Середньому Сході та Європі. Ареали проживання популяцій неандертальців та сучасних людей на деяких територіях накладались один на одного (наприклад на Піренейському півострові та на Середньому Сході) і популяції сучасних людей, які переселялись (також відомих як "кроманьйонці" або європейська рання сучасна людина) до певної міри зхрещувались з ними. Таке змішування може бути причиною генів неандертальців у мешканців Євразії та Океанії часів палеоліту та сучасності.
Важливою відмінністю між Європою та іншими частинами населеного світу була північна широта; археологічні свідчення дозволяють припустити, що людина, неандертальці або кроманьйонці, досягли стоянок у Арктичній Росії вже 40 тисяч років тому.[63]
Першими анатомічно сучасними людьми в Європі вважаються кроманьйонці; вони ввійшли у Євразію через гори Загрос (поблизу сучасного Ірану і східної Туреччини) бл. 50 тис. років тому. Одна їх група досить швидко заселила прибережні території довкола Індійського океану, а друга мігрувала на північ до степів Центральної Азії.[64] Залишки сучасної людини, які датуються 45-43 тисячі років тому, були знайдені в Італії[65] and Britain,[66] а знайдені в європейській частині російської Арктики - 40 тис. років тому.[67][68]
Людина колонізувала території на захід від Уралу, полюючи на оленів,[69] але стикнулася з викликами адаптації; зимові температури сягали мінус 20−30 °C , а можливості знайти паливо та прихисток були обмежені. Вони подорожувлаи пішки та покладалися на полювання високомобільних стад для отримання їжі. Ці виклики були подолані технологічними інноваціями: виробництвом обягу по фігурі зі шкір хутряного звіра; будівництвом притулків з вогнищаями, які використовували кістки як паливо; і риттям “крижаних сховищ” у вічній мерзлоті для зберігання м'яса та кісток.[69][70]
Секвенування мітохондріальної ДНК двох кроманьйонців з печери Пагліччі в Італії, датованої 24-23 тисячі років тому (Paglicci 52 і 12), ідентифікувало мтДНК як гаплогрупу N, типову для цієї групи.[71]
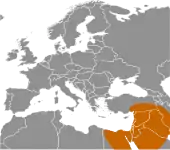
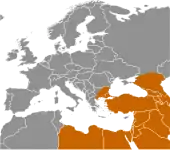
Вважається, що поширення популяції сучасної людини почалось 45 тис. років тому і для колонізації Європи знадобилось 15–20 тис. років.[73][74]
Протягом цього часу неандертальців повільно витісняли. Оскільки заселення Європи зайняло стільки часу, можливо сучасна людина та неандертальців постійно змагались за територію. Неандертальці мали більший мозок та в цілому були більші, з більш важкою статурою, тому ймовірно фізично були сильніші за сучасну Homo sapiens. Оскільки вони прожили в Європі 200 тис. років, вони були більше адаптовані до холодної погоди. Анатомічно сучасна людина, відома як кроманьйонці, з широкою торговою мережею, кращою технологією та тілами, що ймовірно краще були пристосовані до бігу, врешті решт повністю витіснили неандертальців, чиїм останнім притулком став Піренейський півострів. Орієнтовно 25 тис. років тому археологічні свідчення неандертальців припиняються, що свідчить про їх вимирання. Остання відома популяція жила довкола системи печер на віддаленому південному березі Гібралтара між 30 і 24 тисячі років тому.
На основі величини нерівноважного зчеплення вважається, що останній потік генів неандертальців у ранніх предків європейців відбувся 65-47 тисяч років тому. З урахуванням археологічних свідчень та виявлених скамянілостей, цей потік генів ймовірно трапився десь у Західній Євразії, можливо на Середньому Сході.[59] Дослідження показують вищий відсоток генів неандертальців у азійцях, ніж у європейцях.[75][76] Північноафриканські групи поділяють схожий надлишок похідних аллелей з неандертальцями, як і неафриканські популяції, а от африканські групи в Субсахарській Африці є єдиними популяціями сучасних людей, які переважно не мають домішку неандертальців.[77] Пов'язаний з неандертальцями гаплотип B006 гену дистрофіну також був знайденим серед мігруючих груп скотарів у Сахелі та на Сомалійському півострові, які асоціюються з північними популяціями. Відповідно, присутність цього гаплотипу B006 на північному та північносхідному периметрі Африки на південь від Сахари приписується генетичному потоку неафриканського походження.[78]
Східна та Північна Азія
"Тянюанська людина", яка жила на території Китаю бл. 40 тис. років тому, мала суттєвий домішок неандертальців серед предків. Дослідження 2017 року стародавньої ДНК Тянюанськи людини показало, що вона була тісно пов'язана з сучасними популяціями Східної Азії, але не є їх прямими предком.[79] Дослідження 2013 року виявило інтрогресії 18 генів неандертальців у хромосомному регіоні 3p21.31 (регіон HYAL region) у мешканців Східної Азії. Інтрогресійні гаплотипи були позитивно відібрані лише у популяцій Східної Азії, поступово зростаючи починаючи з 45 тис. років до раптового збільшення коефіцієнту рості між 5 і 3,5 тисячами років тому. Они трапляються з високою частотою серед популяцій Східної Азії у порівнянні з іншими євроазійськими популяціями (напр., європейськими та південноазійськими популяціями). Результати дослідження також дозволяють припустити, що ці інтрогресії неандертальців трапилися серед популяції спільних предків мешканців Східної Азії та корінних американців.[80]
Дослідження 2016 року вважає аналіз генетики популяції народу Айни, які мешкають у північній Японії, ключовим для реконструкції раннього заселення Східної Азії. З'ясовано, що Айни представляють більш базову гілку, ніж сучасні популяції фермерів Східної Азії, і ймовірно мають стародавній (до Неоліту) зв'язок з мешканцями північносхідного Сибіру.[81] Дослідження 2013 року пов'язало деякі фенотипні характеристики, які асоціюються з монголоїдною расою, з одиничною мутацією гену EDAR, яка відбулася бл. 35 тис. років тому.[82]
Мітохондріальні гаплогрупи A, B і G виникли бл. 50 тис. років тому, і їх носії відповідно колонізували Сибір, Корею і Японію до 35 тис. років тому. Частини цих популяцій мігрували до Північної Америки під час Останнього льодовикового максимуму.
Євразія
Бл. 20 тис. років тому, орієнтовно через 5 тис. років після вимирання неандертальців, останній льодовиковий максимум змусив мешканців північної півкулі мігрувати до декількох притулків (відомих як рефугіуми) до кінця цього періоду. Вважається, що саме нащадки популяцій, які пережили льодовиковий максимум у цих рефугіумах, потім знову заселили Європу і є предками архаїчних історичних народів. Склад європейських популяцій був потім змінений новими міграціями, в першу чергу внаслідок неолітичної міграції з Середнього Сходу, і пізнішою халколітичними переселеннями, які пов'язують з Індо-Європейською міграцією. Палеолітична археологічна ділянка на ріці Яна, Сибір, на 71° п.ш., лежить значно вище арктичного полярного кола та датується 27 тис. радіовуглецевих років тому, в часи заледеніння. Ділянка демонструє, що люди адаптувалися до цього жорсткого середовища Пізнього Плейстоцену високих широт значно раніше, ніж вважалося.[83]
Припускають, що африканська Кебаранська культура Епіпалеоліту досягла Близького Сходу бл. 18 тис. років тому, принесши туди лук та стріли.[джерело?]
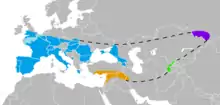
Америки
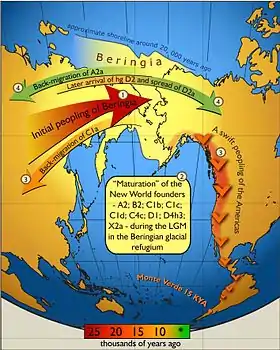
Палеоіндіанці походять від популяцій Центральної Азії, які перейшли суходільний міст Берингії між східним Сибіром та сучасною Аляскою.[84] Люди оселились в Америках до кінця останнього льодовикового періоду, або більш точно останнього льодовикового максимуму, не раніше 23 тис. років тому.[84][85][86][87] Деталі міграцій палеоіндіанців по американському континенту, включно з датами та маршрутами, лишаються предметом дебатів та досліджень.[88]
За традиційною теорією, ці ранні мігранти прийшли, коли рівень моря був значно нижчим з огляду на Четвертинне зледеніння,[85][88] йдучи слідом за стадами вже вимерлої плейстоценової мегафауни по вільних від криги коридорах, які простягались між Лаврентійським і Кордильєрським льодовиковим щитами.[89] За іншим запропонованим маршрутом, пішки або з використанням примітивних човнів, вони мігрували вздовж тихоокеанського узбережжя до Південної Америки (аж до Чилі[90], однак будь-які археологічні докази розселення берегами під час останнього льодовикового періоду зараз приховані декількома сотнями метрів води внаслідок підвищення рівня моря.[91] Нещодавня знахідка генетичних маркерів корінних австралоазійців в Амазонії надала певну підтримку гіпотезі прибережного маршруту.[92][93]
Міграції Голоцену
Початок Голоцену визначають 12 тис. років тому, після завершення Останнього льодовикового максимуму. Під час голоценового кліматичного оптимуму, який розпочався бл. 9 тис. років тому, почалася міграція людських популяцій, які до того були географічно обмежені у рефугіумах. До цього часу більшість частин земної кулі вже були заселені H. sapiens; але значні території, які були вкриті льодовиками, тепер змогли були повтоно заселені.
У цьому періоді відбувається перехід від Мезоліту до Неоліту по всій помірній кліматичній зоні. Неоліт потім поступається Бронзові добі у культурах Старого світу, і поступово бл. 4 тис. років тому виникають історичні записи на Близькому Сході та в Китаї.
Вважається, що масштабні міграції у Мезоліті та Неоліті дали початок досучасному розподілу основних світових мовних сімей, таких як нігеро-конголезькі мови, ніло-сахарські мови, афразійські мови, уральські мови, сино-тибетські мови чи індоєвропейські мови. Спекулятивна ностратична теорія постулює походження основних мовних сімей Євразії (за виключеннями сино-тибетьської) з єдиної протомови, якою розмовляти на початку Голоцену.
Євразія
Оприлюднені 2014 року результати аналізу геному залишків стародавніх людей дозволяють припустити, що сучасне корінне населення Європи значною мірою є нащадками трьох виокремлених родів: "західні мисливці-збирачі", які походять від кроманьйонців Європи, ранні європейські фермери, які прийшли в Європу з Близького Сходу під час неолітичної революції і стародавні північні євразійці, які поширились у Європу в рамках індоєвропейської міграції.[94]
Афроазійську прабатьківщину розміщують у Африці або Азії.
Субсахарська Африка
Вважається, що нілоти виокремились від більш ранньої нерозділеної східносуданської спільноти бл. 5 тис. років тому. Розвиток протонілотів як групи міг бути пов'язаний з їх одомашненням худоби. Східносуданська спільнота мала б існувати значно раніше, ймовірно бл. 7 тис. років тому (а запропонована ніло-сахарська спільнота - у Пізньому палеоліті, бл. 15 тис. років тому). Початковим локусом ранніх носіїв нілотської мови ймовірно була територія на схід від Нілу, у сучасному Південному Судані. Протонілоти 5 тис. років тому переважно займалися випасом худоби, а їх сусіди, прото-центральносуданські народи, переважно вирощували сільськогосподарські культури.[95]
Вважається, що нігеро-конголезька сім'я виникла бл. 6 тис. років тому у Західній чи Центральній Африці. Її поширення могло були повязано з поширенням сахельського сільського господарства у неолітичну добу Африки, після висихання Сахари бл. 39 ст. до н.е..[96] Міграція банту поширила мови Банту у Центральну, Східну та Південну Африку, частково замінивши корінне населення цих регіонів. Почавшись бл. 3 тис. років тому, вона досягла Південної Африки бл. 1 700 років тому.[97]
Тихий океан
.png.webp)
Острови Тихого океану були заселені між бл. 1600 р. до н.е. і 1000 р. н.е. Народ Лапіта, які отримали назву від археологічної ділянки у Лапіта, Нова Каледонія, де їх характерні глиняні вироби були вперше відкриті, був народом, який розмовляв австронезійською та поселився у Ближній Океанії (в першу чергу на архіпелазі Бісмарка у у Папуа-Новій Гвинеї, та на Соломонових островах) бл. 1500 р. до н.е., де відбулося деяке змішування з наявним папуанським населенням. Після того, як вони отримали навички довгих морських подорожей, вони вирушили у 'Віддалену Океанію', ймовірно заселивши Вануату та Нову Каледонію бл. 1200 р. до н.е., потім Фіджі, Самоа та Тонга. На початок 1 тисячоліття до н.е., ця західна частина Полінезії була розкиданою павутиною вдалих популяцій, які оселилися на берегах островів та жили морем. До початку нової ери Мікронезія була повністю заселена; а тропічна східна Полінезія, включно з Таїті, була ймовірно заселена до 700 р.н.е. Останнім регіоном Полінезії, який було досягнуто, ймовірно стала Нова Зеландія, ймовірно десь бл. 1300 р.н.е.[98]
Кариби
Кариби були одним з останніх місць, заселених людиною в Америках. Найстаріші залишки людини відомі з Великих Антильських островів (Куба та Еспаньйола) і датуються між 4000–3500 рр. до н.е.; порівняння у техніці виготовлення знарядь дозволяють припустити, що ці люди прийшли через Юкатанську протоку з Центральної Америки. Існує також багато доказів, що пізніші мігранти десь 2000 року до н.е. і пізніше прийшли з Південної Америки через регіон Оріноко. Нащадки цих мігрантів включають предків народів Таїно іКалінаго (острівні Кариби).[99]
Арктика
Останнім регіоном, який був постійно заселений міграцією людини, стала Арктика.
Найбільш ранніми мешканцями центральної та східної Арктики Півічної Америки вважаються представники так званої Арктичної традиції малих знарядь (AST), яка існувала бл. 2500 років до н.е. AST cскладалася з декількох культур палеоескімосів, включно з культурами Індепенденс та Додорсетською культурою.[100][101]
Інуїти є нащадками культури Туле, яка виникла з західної Аляски бл. 1000 р. н.е. та поступово витіснила Дорсетську культуру.[102][103]
Див. також
- Міграція населення
- Хронологія життя
- Перелік перших людських поселень
- Середній палеоліт
- Хронологія природи
- Хронологія людської еволюції
- Пізній палеоліт
Примітки
- Schlebusch et al., "Southern African ancient genomes estimate modern human divergence to 350,000 to 260,000 years ago", Science, 28 Sep 2017, DOI:10.1126/science.aao6266, Fig. 3
- Архаїчні домішки різного походження відомі у Європі та Азії (неандертальці), Південносхідній Азії та Меланезії (денисівська людина) та у Західній та Південній Африці. Пропорція змінюється в залежності від регіону, але не перевищує 10%: У Євразії вона оцінюється у 1–4% (вища оцінка у 3,4–7,3% дається у праці Lohse, K.; Frantz, L.A.F. (2014). "Neandertal Admixture in Eurasia Confirmed by Maximum-Likelihood Analysis of Three Genomes". Genetics. 196 (4): 1241–1251. DOI:10.1534/genetics.114.162396); у меланезійців оцінюється у 4–6% (Reich et al. (2010). "Genetic history of an archaic hominin group from Denisova Cave in Siberia". Nature. 468 (7327): 1053–1060. DOI:10.1038/nature09710.
- Zhu, Zhaoyu; Dennell, Robin; Huang, Weiwen; Wu, Yi; Qiu, Shifan; Yang, Shixia; Rao, Zhiguo; Hou, Yamei; Xie, Jiubing; Han, Jiangwei; Ouyang, Tingping (2018). Hominin occupation of the Chinese Loess Plateau since about 2.1 million years ago. Nature 559 (7715): 608–612. PMID 29995848. doi:10.1038/s41586-018-0299-4.
- Out of Africa I: The First Hominin Colonization of Eurasia. Vertebrate Paleobiology and Paleoanthropology. Springer. 2010. ISBN 978-90-481-9035-5.
- Garcia, T.; Féraud, G.; Falguères, C.; de Lumley, H.; Perrenoud, C.; Lordkipanidze, D. (2010). Earliest human remains in Eurasia: New 40Ar/39Ar dating of the Dmanisi hominid-bearing levels, Georgia. Quaternary Geochronology 5 (4): 443–451. doi:10.1016/j.quageo.2009.09.012.
- R. Zhu et al. (2004), New evidence on the earliest human presence at high northern latitudes in northeast Asia.
- Rixiang Zhu; Zhisheng An; Richard Pott; Kenneth A. Hoffman (June 2003). Magnetostratigraphic dating of early humans in China. Earth-Science Reviews 61 (3–4): 191–361. Bibcode:2003ESRv...61..191A. doi:10.1016/S0012-8252(02)00110-1. Архів оригіналу за 24 липня 2011.
- Hopkin M (26 березня 2008). 'Fossil find is oldest European yet'. Nature News. doi:10.1038/news.2008.691.
- Bednarik RG (2003). Seafaring in the Pleistocene. Cambridge Archaeological Journal 13 (1): 41–66. doi:10.1017/S0959774303000039.
ScienceNews summary - Homo erectus soloensis, знайдений на острові Ява, вважається найбільш пізнім відомими представником H. erectus. Спочатку його датували 50-40 тис. років тому, але дослідження 2011 року відсунуло дату вимирання H. e. soloensis принаймні до 143 тис. років тому, а ймовірно і раніше 550 тис. років. Див. Indriati E, Swisher CC III, Lepre C, Quinn RL, Suriyanto RA, et al. 2011 The Age of the 20 Meter Solo River Terrace, Java, Indonesia and the Survival of Homo erectus in Asia. PLoS ONE 6(6): e21562. DOI:10.1371/journal.pone.0021562.
- Ewen Callaway An Arctic refuge for Neanderthals? 12 May 2011 (nature.com).
- Xu, D. (2017). Archaic Hominin Introgression in Africa Contributes to Functional Salivary MUC7 Genetic Variation. Molecular Biology and Evolution 34 (10): 2704–2715. PMC 5850612. PMID 28957509. doi:10.1093/molbev/msx206.
- Hammer, M.F.; Woerner, A.E.; Mendez, F.L.; Watkins, J.C.; Wall, J.D. (2011). Genetic evidence for archaic admixture in Africa. Proceedings of the National Academy of Sciences 108 (37): 15123–15128. Bibcode:2011PNAS..10815123H. PMC 3174671. PMID 21896735. doi:10.1073/pnas.1109300108.
- Callaway, E. (26 липня 2012). Hunter-gatherer genomes a trove of genetic diversity. Nature. doi:10.1038/nature.2012.11076.
- Lachance, J.; Vernot, B.; Elbers, C.C.; Ferwerda, B.; Froment, A.; Bodo, J.M. (2012). Evolutionary History and Adaptation from High-Coverage Whole-Genome Sequences of Diverse African Hunter-Gatherers. Cell 150 (3): 457–469. PMC 3426505. PMID 22840920. doi:10.1016/j.cell.2012.07.009.
- Dirks, Paul HGM; Roberts, Eric M.; Hilbert-Wolf, Hannah; Kramers, Jan D.; Hawks, John; Dosseto, Anthony; Duval, Mathieu; Elliott, Marina; Evans, Mary; Grün, Rainer; Hellstrom, John; Herries, Andy IR; Joannes-Boyau, Renaud; Makhubela, Tebogo V.; Placzek, Christa J.; Robbins, Jessie; Spandler, Carl; Wiersma, Jelle; Woodhead, Jon; Berger, Lee R. (9 травня 2017). The age of Homo naledi and associated sediments in the Rising Star Cave, South Africa. eLife 6. PMC 5423772. PMID 28483040. doi:10.7554/eLife.24231.
- Lopez, Saioa; van Dorp, Lucy; Hellenthal, Garrett (2016). Human Dispersal Out of Africa: A Lasting Debate. Evolutionary Bioinformatics 11s2 (Suppl 2): 57–68. ISSN 1176-9343. PMC 4844272. PMID 27127403. doi:10.4137/EBO.S33489.
- Callaway, Ewen (2016). Oldest ancient-human DNA details dawn of Neanderthals. Nature 531 (7594): 296–286. Bibcode:2016Natur.531..296C. PMID 26983523. doi:10.1038/531286a.
- David Richter (8 червня 2017). The age of the hominin fossils from Jebel Irhoud, Morocco, and the origins of the Middle Stone Age. Nature 546 (7657): 293–296. Bibcode:2017Natur.546..293R. PMID 28593967. doi:10.1038/nature22335. "Тут ми повідомляємо про вік, визначених термолюмінісцентним датуванням нагрітих вогнем артефактів з кремнію, які були знайдені при розкопках археологічної ділянки Середньої Кам'яної доби у Джебель-Ірхуд, Марокко, і які прямо пов'язуються зі щойно виявленими залишками H. sapiens. Середньозважений вік цих артефактів та скамянілостей Середньої Кам'яної доби визначено на рівні 315 ± 34 тис. років тому. Це значення підтримується перерахованими урановими серіями з резонансом спіну електрону, які дають вік 286 ± 32 тис.років для зубу решток гомініну Irhoud 3."; Smith TM, Tafforeau P, Reid DJ, etal (April 2007). Earliest evidence of modern human life history in North African early Homo sapiens. Proceedings of the National Academy of Sciences of the United States of America 104 (15): 6128–33. Bibcode:2007PNAS..104.6128S. PMC 1828706. PMID 17372199. doi:10.1073/pnas.0700747104.
- Mcdougall, I.; Brown, H.; Fleagle, G. (Feb 2005). Stratigraphic placement and age of modern humans from Kibish, Ethiopia. Nature 433 (7027): 733–736. Bibcode:2005Natur.433..733M. ISSN 0028-0836. PMID 15716951. doi:10.1038/nature03258.
- Є пропозиція датувати присутність ранньої сучасної людини за межами Африки аж до 177 тис. років тому. Див. Hershkovitz et al., "The earliest modern humans outside Africa", Science 26 Jan 2018: Vol. 359, Issue 6374, pp. 456–459 DOI:10.1126/science.aap8369
- Eleanor Scerri, "The Stone Age Archaeology of West Africa", African History, 2017, DOI:10.1093/acrefore/9780190277734.013.137.
- Schlebusch et al., "Southern African ancient genomes estimate modern human divergence to 350,000 to 260,000 years ago" Science, 28 Sep 2017, DOI:10.1126/science.aao6266. Estimated split times given in the source cited (in kya): Human-Neanderthal: 530–690, Deep Human [H. sapiens]: 250–360, NKSP-SKSP: 150–190, Out of Africa (OOA): 70–120.
- Rito T, Richards MB, Fernandes V, Alshamali F, Cerny V, Pereira L, Soares P., "The first modern human dispersals across Africa", PLoS One 2013 Nov 13; 8(11):e80031. DOI:10.1371/journal.pone.0080031. "By ~130 ka two distinct groups of anatomically modern humans co-existed in Africa: broadly, the ancestors of many modern-day Khoe and San populations in the south and a second central/eastern African group that includes the ancestors of most extant worldwide populations. Early modern human dispersals correlate with climate changes, particularly the tropical African “megadroughts” of MIS 5 (marine isotope stage 5, 135–75 ka) which paradoxically may have facilitated expansions in central and eastern Africa, ultimately triggering the dispersal out of Africa of people carrying haplogroup L3 ~60 ka. Two south to east migrations are discernible within haplogroup L0. One, between 120 and 75 ka, represents the first unambiguous long-range modern human dispersal detected by mtDNA and might have allowed the dispersal of several markers of modernity. A second one, within the last 20 ka signalled by L0d, may have been responsible for the spread of southern click-consonant languages to eastern Africa, contrary to the view that these eastern examples constitute relicts of an ancient, much wider distribution."
- Jarvis JP, Scheinfeldt LB, Soi S, Lambert C, Omberg L, Ferwerda B, et al. (2012) Patterns of Ancestry, Signatures of Natural Selection, and Genetic Association with Stature in Western African Pygmies. PLoS Genet 8(4): e1002641. DOI:10.1371/journal.pgen.1002641. López Herráez D, Bauchet M, Tang K, Theunert C, Pugach I, Li J, et al. (2009) Genetic Variation and Recent Positive Selection in Worldwide Human Populations: Evidence from Nearly 1 Million SNPs. PLoS ONE 4(11): e7888. DOI:10.1371/journal.pone.0007888. Tishkoff SA, Reed FA, Friedlaender FR, Ehret C, Ranciaro A, Froment A, Hirbo JB, Awomoyi AA, Bodo JM, Doumbo O, Ibrahim M, Juma AT, Kotze MJ, Lema G, Moore JH, Mortensen H, Nyambo TB, Omar SA, Powell K, Pretorius GS, Smith MW, Thera MA, Wambebe C, Weber JL, Williams SM (2009). The Genetic Structure and History of Africans and African Americans. Science 324 (5930): 1035–1044. Bibcode:2009Sci...324.1035T. PMC 2947357. PMID 19407144. doi:10.1126/science.1172257. (Supplementary Data) Quintana-Murci et al. 2008. Maternal traces of deep common ancestry and asymmetric gene flow between Pygmy hunter–gatherers and Bantu-speaking farmers 'Proceedings of the National Academy of Sciences of the United States of America'. 105(5): 1599, fig. 3. Patin E, Laval G, Barreiro LB, Salas A, Semino O, Santachiara-Benerecetti S, Kidd KK, Kidd JR, Van der Veen L, Hombert JM, Gessain A, Froment A, Bahuchet S, Heyer E, Quintana-Murci L (2009). Inferring the Demographic History of African Farmers and Pygmy Hunter–Gatherers Using a Multilocus Resequencing Data Set. PLoS Genetics 5 (4): e1000448. PMC 2661362. PMID 19360089. doi:10.1371/journal.pgen.1000448. "We studied the branching history of Pygmy hunter–gatherers and agricultural populations from Africa and estimated separation times and gene flow between these populations. The model identified included the early divergence of the ancestors of Pygmy hunter–gatherers and farming populations ~60,000 years ago, followed by a split of the Pygmies' ancestors into the Western and Eastern Pygmy groups ~20,000 years ago."
- Holen, Steven R. (2017). A 130,000-year-old archaeological site in southern California, USA. Nature 544 (7651): 479–483. Bibcode:2017Natur.544..479H. PMID 28447646. doi:10.1038/nature22065.
- Lawler, Andrew (2011). Did Modern Humans Travel Out of Africa Via Arabia?. Science 331 (6016): 387. Bibcode:2011Sci...331..387L. PMID 21273459. doi:10.1126/science.331.6016.387.
- Trail of 'Stone Breadcrumbs' Reveals the Identity of One of the First Human Groups to Leave Africa ScienceDaily (Nov. 30, 2011)
- Ознаки більш раннього виходу людини з Африки
- Liu, Wu (2010). Human remains from Zhirendong, South China, and modern human emergence in East Asia. Proceedings of the National Academy of Sciences 107 (45): 19201–19206. Bibcode:2010PNAS..10719201L. PMC 2984215. PMID 20974952. doi:10.1073/pnas.1014386107. (the authors seem to accept that the individual has African recent ascentry, but with Asian archaic human admixture). Див. також Robin Dennell, Two interpretations of the Zhirendong mandible described by Liu and colleagues, Nature Volume: 468, 25 November 2010, pages: 512–513, DOI:10.1038/468512a; Brief comments at Modern Humans Emerged Far Earlier Than Previously Thought, Fossils from China Suggest, ScienceDaily (Oct. 25, 2010) https://www.sciencedaily.com/releases/2010/10/101025172924.htm; and Oldest Modern Human Outside of Africa Found: http://news.nationalgeographic.com/news/2010/10/101025-oldest-human-fossil-china-out-of-africa-science/
- Shena, Guanjun (2002). U-Series dating of Liujiang hominid site in Guangxi, Southern China. Journal of Human Evolution 43 (6): 817–829. PMID 12473485. doi:10.1006/jhev.2002.0601.
- Lunadong fossils support theory of earlier dispersal of modern man
- Callaway, Ewen (2015). Teeth from China reveal early human trek out of Africa. Nature. doi:10.1038/nature.2015.18566. Процитовано 23 жовтня 2015.
- Balter, Michael (2010). Of Two Minds About Toba's Impact. Science 327 (5970): 1187–1188. Bibcode:2010Sci...327.1187B. PMID 20203021. doi:10.1126/science.327.5970.1187-a.
- Vicente M. Cabrera (2017). Carriers of mitochondrial DNA macrohaplogroup L3 basic lineages migrated back to Africa from Asia around 70,000 years ago. Проігноровано невідомий параметр
|biorxiv=(довідка) - Zhivotovsky; Rosenberg, NA; Feldman, MW (2003). Features of Evolution and Expansion of Modern Humans, Inferred from Genomewide Microsatellite Markers. American Journal of Human Genetics 72 (5): 1171–1186. PMC 1180270. PMID 12690579. doi:10.1086/375120.
- Stix, Gary (2008). The Migration History of Humans: DNA Study Traces Human Origins Across the Continents. Процитовано 14 червня 2011.
- Armitage, Simon J.; Jasim, Sabah A.; Marks, Anthony E.; Parker, Adrian G.; Usik, Vitaly I.; Uerpmann, Hans-Peter (2011). Hints Of Earlier Human Exit From Africa. Science 331 (6016): 453–456. Bibcode:2011Sci...331..453A. PMID 21273486. doi:10.1126/science.1199113. Процитовано 1 травня 2011.
- Scientists discover oldest known modern human fossil outside of Africa: Analysis of fossil suggests Homo sapiens left Africa at least 50,000 years earlier than previously thought. ScienceDaily (англ.). Процитовано 27 січня 2018.
- Ghosh, Pallab (2018). Modern humans left Africa much earlier. BBC News (en-GB). Процитовано 27 січня 2018.
- Dennell, Robin; Petraglia, Michael D. (2012). The dispersal of Homo sapiens across southern Asia: how early, how often, how complex?. Quaternary Science Reviews 47: 15–22. Bibcode:2012QSRv...47...15D. doi:10.1016/j.quascirev.2012.05.002.
- Prüfer, Kay (2013). The complete genome sequence of a Neanderthal from the Altai Mountains. Nature 505 (7481): 43–49. Bibcode:2014Natur.505...43P. PMC 4031459. PMID 24352235. doi:10.1038/nature12886.
- Bowler, James M. (2003). New ages for human occupation and climatic change at Lake Mungo, Australia. Nature 421 (6925): 837–840. Bibcode:2003Natur.421..837B. PMID 12594511. doi:10.1038/nature01383.
- Callaway, Ewen (22 вересня 2011). First Aboriginal genome sequenced. Nature. doi:10.1038/news.2011.551.
- Reich (2011). Denisova Admixture and the First Modern Human Dispersals into Southeast Asia and Oceania. The American Journal of Human Genetics 89 (4): 516–528. PMC 3188841. PMID 21944045. doi:10.1016/j.ajhg.2011.09.005.
- Choi, Charles (22 вересня 2011). Now-Extinct Relative Had Sex with Humans Far and Wide. LiveScience.
- Cooper A.; Stringer C.B. (2013). Did the Denisovans Cross the Wallace Line. Science 342 (6156): 321–333. Bibcode:2013Sci...342..321C. PMID 24136958. doi:10.1126/science.1244869.
- Salleh, Anna (18 жовтня 2013). Humans dated ancient Denisovan relatives beyond the Wallace Line. ABC Science.
- First Mariners – Archaeology Magazine Archive. Archive.archaeology.org. Retrieved on 2013-11-16.
- Pleistocene Sea Level Maps. Fieldmuseum.org. Процитовано 23 вересня 2010.
- Rasmussen, M (Oct 2011). An Aboriginal Australian genome reveals separate human dispersals into Asia. Science 334 (6052): 94–98. Bibcode:2011Sci...334...94R. PMC 3991479. PMID 21940856. doi:10.1126/science.1211177.
- Hudjashov G, Kivisild T, Underhill PA (May 2007). Revealing the prehistoric settlement of Australia by Y chromosome and mtDNA analysis. Proc Natl Acad Sci USA 104 (21): 8726–8730. Bibcode:2007PNAS..104.8726H. PMC 1885570. PMID 17496137. doi:10.1073/pnas.0702928104.
- Wade, Nicholas (8 травня 2007). From DNA Analysis, Clues to a Single Australian Migration. Australia: Nytimes.com. Процитовано 1 травня 2011.
- Clarkson, Chris; Smith, Mike; Marwick, Ben; Fullagar, Richard; Wallis, Lynley A.; Faulkner, Patrick; Manne, Tiina; Hayes, Elspeth; Roberts, Richard G.; Jacobs, Zenobia; Carah, Xavier; Lowe, Kelsey M.; Matthews, Jacqueline; Florin, S. Anna (June 2015). The archaeology, chronology and stratigraphy of Madjedbebe (Malakunanja II): A site in northern Australia with early occupation. Journal of Human Evolution 83: 46–64. PMID 25957653. doi:10.1016/j.jhevol.2015.03.014. Проігноровано невідомий параметр
|hdl=(довідка) - Clarkson, Chris; Jacobs, Zenobia; Marwick, Ben; Fullagar, Richard; Wallis, Lynley; Smith, Mike; Roberts, Richard G.; Hayes, Elspeth; Lowe, Kelsey; Carah, Xavier; Florin, S. Anna; McNeil, Jessica; Cox, Delyth; Arnold, Lee J.; Hua, Quan; Huntley, Jillian; Brand, Helen E. A.; Manne, Tiina; Fairbairn, Andrew; Shulmeister, James; Lyle, Lindsey; Salinas, Makiah; Page, Mara; Connell, Kate; Park, Gayoung; Norman, Kasih; Murphy, Tessa; Pardoe, Colin (19 липня 2017). Human occupation of northern Australia by 65,000 years ago. Nature 547 (7663): 306–310. Bibcode:2017Natur.547..306C. PMID 28726833. doi:10.1038/nature22968.
- Flannery, Tim (2002), "The Future Eaters: An Ecological History of the Australasian Lands and People" (Grove Press)
- Mellars, Paul (11 серпня 2006). Going East: New Genetic and Archaeological Perspectives on the Modern Human Colonization of Eurasia. Science 313 (5788): 796–800. Bibcode:2006Sci...313..796M. PMID 16902130. doi:10.1126/science.1128402.
- Fossil Teeth Put Humans in Europe Earlier Than Thought. The New York Times. 2 листопада 2011.
- Sankararaman, S.; Patterson, N.; Li, H.; Pääbo, S.; Reich, D; Akey, J.M. (2012). The Date of Interbreeding between Neandertals and Modern Humans. PLoS Genetics 8 (10): e1002947. PMC 3464203. PMID 23055938. doi:10.1371/journal.pgen.1002947.
- Harpending, Henry; Cochran, Gregory (2009). The 10,000 Year Explosion. Basic Books. с. 214. ISBN 978-0-465-00221-4. Процитовано 1 грудня 2015.
- Ambrose, Stanley (1998). Late Pleistocene human population bottlenecks, volcanic winter, and differentiation of modern humans. Journal of Human Evolution 34 (6): 623–651. PMID 9650103. doi:10.1006/jhev.1998.0219.
- Oppenheimer, Stephen "Out of Eden: Peopling of the World" (Robinson; New Ed edition (March 1, 2012))
- Pavlov, Pavel; John Inge Svendsen; Svein Indrelid (6 вересня 2001). Human presence in the European Arctic nearly 40,000 years ago. Nature 413 (6851): 64–67. Bibcode:2001Natur.413...64P. PMID 11544525. doi:10.1038/35092552.
- Atlas of human journey: 45–40,000. Genographic Project. National Geographic Society. 1996–2010.
- Benazzi, S.; Douka, K.; Fornai, C.; Bauer, C.C.; Kullmer, O.; Svoboda, J.F.; Pap, I.; Mallegni, F.; Bayle, P.; Coquerelle, M.; Condemi, S.; Ronchitelli, A.; Harvati, K.; Weber, G.W. (2011). Early dispersal of modern humans in Europe and implications for Neanderthal behaviour. Nature 479 (7374): 525–528. Bibcode:2011Natur.479..525B. PMID 22048311. doi:10.1038/nature10617.
- Higham, T.; Compton, T.; Stringer, C.; Jacobi, R.; Shapiro, B.; Trinkaus, E.; Chandler, B.; Gröning, F.; Collins, C.; Hillson, S.; o’Higgins, P.; Fitzgerald, C.; Fagan, M. (2011). The earliest evidence for anatomically modern humans in northwestern Europe. Nature 479 (7374): 521–524. Bibcode:2011Natur.479..521H. PMID 22048314. doi:10.1038/nature10484.
- Pavlov, Pavel; Svendsen, John Inge; Indrelid, Svein (2001). Human presence in the European Arctic nearly 40,000 years ago. Nature 413 (6851): 64–67. PMID 11544525. doi:10.1038/35092552.
- Mamontovaya Kurya:an enigmatic, nearly 40000 years old Paleolithic site in the Russian Arctic.
- Hoffecker, J. (2006). A Prehistory of the North: Human Settlements of the Higher Latitudes. New Jersey: Rutgers University Press. с. 101.
- Hoffecker, John F. (2002). Desolate landscapes: Ice-Age settlement in Eastern Europe. New Brunswick: Rutgers University Press. с. 158–162, 217–233.
- Caramelli, D; Lalueza-Fox, C; Vernesi, C; Lari, M; Casoli, A; Mallegni, F; Chiarelli, B; Dupanloup, I; Bertranpetit, J; Barbujani, G; Bertorelle, G (May 2003). Evidence for a genetic discontinuity between Neandertals and 24,000-year-old anatomically modern Europeans. Proceedings of the National Academy of Sciences of the United States of America 100 (11): 6593–6597. Bibcode:2003PNAS..100.6593C. ISSN 0027-8424. PMC 164492. PMID 12743370. doi:10.1073/pnas.1130343100.
- Currat M, Excoffier L (2004). Modern Humans Did Not Admix with Neanderthals during Their Range Expansion into Europe. PLoS Biol 2 (12): e421. PMC 532389. PMID 15562317. doi:10.1371/journal.pbio.0020421.
- Maca-Meyer N, González AM, Larruga JM, Flores C, Cabrera VM (2001). Major genomic mitochondrial lineages delineate early human expansions. BMC Genet. 2 (1): 13. PMC 55343. PMID 11553319. doi:10.1186/1471-2156-2-13.
- Currat M, Excoffier L (Dec 2004). Modern humans did not admix with Neanderthals during their range expansion into Europe. PLoS Biol. 2 (12): e421. PMC 532389. PMID 15562317. doi:10.1371/journal.pbio.0020421.
- Meyer, M.; Kircher, M.; Gansauge, M.T.; Li, H.; Racimo, F.; Mallick, S. (2012). A High-Coverage Genome Sequence from an Archaic Denisovan Individual. Science 338 (6104): 222–226. Bibcode:2012Sci...338..222M. PMC 3617501. PMID 22936568. doi:10.1126/science.1224344.
- Wall, J.D.; Yang, M.A.; Jay, F.; Kim, S.K.; Durand, E.Y.; Stevison, L.S. (2013). Higher Levels of Neanderthal Ancestry in East Asians than in Europeans. Genetics 194 (1): 199–209. PMC 3632468. PMID 23410836. doi:10.1534/genetics.112.148213.
- Sánchez-Quinto, F.; Botigué, L.R.; Civit, S.; Arenas, C.; Ávila-Arcos, M.C.; Bustamante, C.D. (2012). North African Populations Carry the Signature of Admixture with Neandertals. PLoS ONE 7 (10): e47765. Bibcode:2012PLoSO...747765S. PMC 3474783. PMID 23082212. doi:10.1371/journal.pone.0047765. «We found that North African populations have a significant excess of derived alleles shared with Neandertals, when compared to sub-Saharan Africans. This excess is similar to that found in non-African humans, a fact that can be interpreted as a sign of Neandertal admixture. Furthermore, the Neandertal's genetic signal is higher in populations with a local, pre-Neolithic North African ancestry. Therefore, the detected ancient admixture is not due to recent Near Eastern or European migrations. Sub-Saharan populations are the only ones not affected by the admixture event with Neandertals.»
- Yotova, Vania (2011). An X-linked haplotype of Neandertal origin is present among all non-African populations. Molecular Biology and Evolution 28 (7): 1957–1962. PMID 21266489. doi:10.1093/molbev/msr024. «Of 1,420 sub-Saharan chromosomes, only one copy of B006 was observed in Ethiopia, and five in Burkina Faso, one among the Rimaibe and four among the Fulani and Tuareg, nomad-pastoralists known for having contacts with northern populations (supplementary table S1, Supplementary Material online). B006 only occurrence at the northern and northeastern outskirts of sub-Saharan Africa is thus likely to be a result of gene flow from a non-African source.»
- Yang et al., "40,000-Year-Old Individual from Asia Provides Insight into Early Population Structure in Eurasia", Current Biology Volume 27, Issue 20, pp. 3202–3208.e9, 23 October 2017, DOI:10.1016/j.cub.2017.09.030.
- Ding, Q.; Hu, Y.; Xu, S.; Wang, J.; Jin, L. (2014) [Online 2013]. Neanderthal Introgression at Chromosome 3p21.31 was Under Positive Natural Selection in East Asians. Molecular Biology and Evolution 31 (3): 683–695. PMID 24336922. doi:10.1093/molbev/mst260..
- Jeong et al., "Deep History of East Asian Populations Revealed Through Genetic Analysis of the Ainu", Genetics. 2016 Jan;202(1):261–272. DOI:10.1534/genetics.115.178673.
- Впливу мутації зазнали такі характеристики як потові залози, товщина волосся та тканини грудей. Kamberov et al., "Modeling Recent Human Evolution in Mice by Expression of a Selected EDAR Variant", Cell Volume 152, Issue 4, pp. 691–702, 14 February 2013, DOI:10.1016/j.cell.2013.01.016. Звіт журналістів: East Asian Physical Traits Linked to 35,000-Year-Old Mutation, NYT, 14 February 2013. Генетика Східної Азії демонструє ряд концентрованих аллелей, що свідчить про тиски відбору; це зокрема стосується таких генів як EDAR, ADH1B, ABCC1 і ALDH2. Східноазійські типи ADH1B асоціюються з одомашненням рису і тому виникли не раніше бл. 11 тис. років тому. Peng, Y. et al. The ADH1B Arg47His polymorphism in East Asian populations and expansion of rice domestication in history. BMC Evolutionary Biology 10, 15 (2010).
- Pitulko, V.V.; Nikolsky, P.A.; Girya, E.Y.; Basilyan, A.E.; Tumskoy, V.E.; Koulakov, S.A.; Astakhov, S.N.; Pavlova, E.Y. та ін. (2004). The Yana RHS Site: Humans in the Arctic Before the Last Glacial Maximum. Science 303 (5654): 52–56. Bibcode:2004Sci...303...52P. PMID 14704419. doi:10.1126/science.1085219.
- Wells, Spencer; Read, Mark (2002). The Journey of Man – A Genetic Odyssey (Digitised online by Google books). Random House. с. 138–140. ISBN 978-0-8129-7146-0. Процитовано 21 листопада 2009.
- Fitzhugh, Drs.William; Goddard, Ives; Ousley, Steve; Owsley, Doug; Stanford., Dennis. Paleoamerican. Smithsonian Institution Anthropology Outreach Office. Архів оригіналу за 5 січня 2009. Процитовано 15 січня 2009.
- A DNA Search for the First Americans Links Amazon Groups to Indigenous Australians. Smithsonian.com. Smithsonian Institution. Процитовано 28 серпня 2015.
- Bonatto, S. L.; Salzano, F. M. (1997). A single and early migration for the peopling of greater America supported by mitochondrial DNA sequence data. Proceedings of the National Academy of Sciences 94 (5): 1866–1871. Bibcode:1997PNAS...94.1866B. PMC 20009. PMID 9050871. doi:10.1073/pnas.94.5.1866.
- Atlas of the Human Journey-The Genographic Project. National Geographic Society. 1996–2008. Архів оригіналу за 1 травня 2011. Процитовано 27 січня 2017.
- The peopling of the Americas: Genetic ancestry influences health. Scientific American. Процитовано 17 листопада 2009.
- Fladmark, K. R. (1979). Alternate Migration Corridors for Early Man in North America. American Antiquity, Vol. 44, No. 1 (Jan., 1979), P2 44 (1): 55–69. JSTOR 279189.[недоступне посилання з 01.09.2015]
- 68 Responses to "Sea will rise 'to levels of last Ice Age'". Center for Climate Systems Research, Columbia University. Процитовано 17 листопада 2009.
- accessed 26th October 2015 Genetic studies link indigenous peoples in the Amazon and Australasia Science Daily, July 21, 2015.
- Callaway, Ewen (2015). 'Ghost population' hints at long-lost migration to the Americas: Present-day Amazonians share an unexpected genetic link with Asian islanders, hinting at an ancient trek. Nature News. doi:10.1038/nature.2015.18029.
- Gibbons, Ann (4 вересня 2014). Three-part ancestry for Europeans. Science. American Association for the Advancement of Science. Архів оригіналу за 11 жовтня 2014. Процитовано 30 жовтня 2014.
- John Desmond Clark, From Hunters to Farmers: The Causes and Consequences of Food Production in Africa, University of California Press, 1984, p. 31
- Igor Kopytoff, The African Frontier: The Reproduction of Traditional African Societies (1989), 9–10 (cited afer Igbo Language Roots and (Pre)-History, A Mighty Tree, 2011).
- Vansina, J. (1995). New Linguistic Evidence and 'The Bantu Expansion'. Journal of African History 36 (2): 173–195. JSTOR 182309. doi:10.1017/S0021853700034101.
- Pacific migrations – Te Ara Encyclopedia of New Zealand.
- Fagan, B.M. (2007). People of the earth: An introduction to world prehistory. Upper Saddle River, NJ: Pearson Prentice Hall
- Hoffecker, John F. (2005). A prehistory of the north: human settlement of the higher latitudes. Rutgers University Press. с. 130. ISBN 978-0-8135-3469-5.
- Gibbon, pp. 28–31
- Rigby, Bruce. 101. Qaummaarviit Historic Park, Nunavut Handbook. Архів оригіналу за 29 травня 2006. Процитовано 2 жовтня 2009.
- Wood, Shannon Raye (April 1992). Tooth Wear and the Sexual Division of Labour in an Inuit Population. Department of Archaeology University of Saskatchewan. Simon Fraser University. Процитовано 2 жовтня 2009.
{kind=link}
Подальше читання
- Demeter F, Shackelford LL, Bacon AM, Duringer P, Westaway K, Sayavongkhamdy T, Braga J, Sichanthongtip P, Khamdalavong P, Ponche JL (2012). Anatomically modern human in Southeast Asia (Laos). Proceedings of the National Academy of Sciences 109 (36): 14375–14380. Bibcode:2012PNAS..10914375D. PMC 3437904. PMID 22908291. doi:10.1073/pnas.1208104109.
- Jared Diamond, Guns, germs and steel. A short history of everybody for the last 13'000 years, 1997.
- Reich, David (2018). Who We Are And How We Got Here – Ancient DNA and the New Science of the Human Past. Pantheon Books. ISBN 978-1-101-87032-7.
- Diamond, Jared (20 квітня 2018). A Brand-New Version of Our Origin Story. The New York Times. Процитовано 23 квітня 2018.
- Veeramah, Krishna R.; Hammer, Michael F. (4 лютого 2014). The impact of whole-genome sequencing on the reconstruction of human population history. Nature Reviews Genetics 15 (3): 149–162. PMID 24492235. doi:10.1038/nrg3625.