Фоторецепторна оптика
Фоторецепторна оптика - це наука, яка вивчає вплив оптичних властивостей фоторецепторів сітківки (їх структури, розташування, орієнтації, форми, показник заломлення) на схоплення світла, його поширення і поглинання зоровим пігментом та перетворення у вихідний електричний сигнал [1] [2].
Фоторецептор у сітківці виконує подвійне завдання: спрямовує світло до зорових пігментів і використовує пігмент для перетворення енергії світла в нейронний сигнал. Відтак, фоторецептор виконує як оптичні, так і фотохімічні функції.
Фоторецепторна оптика зародилась в 1860—70-х рр., коли був відкритий зоровий пігмент (фотопігмент) у зовнішньому сегменті фоторецептора і виявлено, що фоторецепторні органели мають вищий показник заломлення, аніж середовище довкола фоторецепторів [3]. Внаслідок вищого показника заломлення та маючи діаметр, близький до довжини хвилі видимого світла, фоторецептор хребетних та безхребетних діє, як оптичний хвилевід, ефективно спрямовуючи падаюче світло до місцезнаходження зорового пігменту [2].
У сітківці хребетних присутні два класи фоторецепторів – палички і колбочки, які відповідають за нічний, безколірний та денний, кольоровий зір, відповідно[4]. Колбочкові фоторецептори сітківки спрямовані приблизно до центру зіниці ока та мають обмежений кут прийняття променів падаючого світла[5] . Внаслідок цього відносна ефективність світлових променів, що досягають фоторецептора, є максимальною для «головного світлового променя» (відомого як b-промінь, який з'єднує центр зіниці із центром хвилеводу) та спадає для «периферійних променів» (a-променів; вищі апертурні кути). Цей феномен лежить в основі ефекту Стайлса-Кроуфорда, що є свого роду аподизацією зіниці[6]. Така зменшена світлова ефективність периферійних світлових променів послаблює їх променеву аберацію та внутріочне розсіяння світла, уможливлюючи більш ефективне схоплення світла фоторецепторними колбочками в центральній ямці сітківки і, як наслідок, вищу гостроту та роздільну здатність зору[7].
Фоторецепторна оптика, як наука, намагається пояснити структурну основу абсолютної, спектральної, напрямкової та поляризаційної чутливості фоторецептора сітківки [3].
Оптика біологічних тканин
Оптичні властивості фоторецепторів невід'ємно пов'язані із оптикою ока (рогівкою, кришталиком, зіницею, скловидним тілом) і перевернутою архітектурою сітківки, що визначають поведінку та величину падаючого світла, яке потрапляє до фоторецептора. Тому для розуміння природи проходження і поглинання світла в фоторецепторі, фоторецепторна оптика бере до уваги оптичні властивості ока та сітківки, що є подібними до інших біологічних тканин, але також характеризуються унікальними рисами, які й обумовлюють ефективність внутріочного проходження світла і його поглинання фоторецепторами.
Перш ніж розглянути оптичні властивості ока і сітківки хребетних і їх вплив на фоторецепторну оптику, слід в загальних рисах охарактеризувати оптичні властивості біологічних тканин, які мають спільні риси з оптичними властивостями ока. Тому теорія взаємодії світла з біологічними тканинами використовується для опису та моделювання оптики ока, сітківки і фоторецепторів.
Оптична неоднорідність біологічних тканин
Оптичні властивості будь-якої біологічної тканини характеризуються її показником заломлення, шорсткістю поверхні та ступенем однорідності [8].
При проходженні світла через середовище відбувається уповільнення його швидкості, що залежить від оптичної густини даного середовища. Подібне зменшення швидкості світла в середовищі використовується для визначення його показника заломлення, n – відношення фазової швидкості світла у вакуумі c до швидкості світла в даному середовищі, v: [9].

Показник заломлення є фундаментальною фізичною величиною, що зазвичай виражається, як корінь з добутку діелектричної проникності з магнітною проникністю :



При проходженні світла через середовище, певна його частка загасає (внаслідок поглинання), що виражається через комплексний показник заломлення:

де дійсна частина, n - це показник заломлення і описує фазову швидкість світла, тоді як уявна частина, k (відома, як коефіцієнт екстинкції), описує величину загасання світла в середовищі[9]. Клітинні органели та мембрани є структурно неоднорідними, що спричиняє і їх оптичну неоднорідність, а саме відмінні, нерегулярно розподілені показники заломлення (до прикладу, водний цитозоль має меншу оптичну густину, аніж мітохондрія) [8]. Тому, з оптичної точки зору, всі біологічні клітини є фазовими об’єктами: при взаємодії світла з клітинними неоднорідностями, що мають вищий показник заломлення, аніж оточуюче середовище, відбувається зсув фази світла, Δφ , що може бути виявлений з допомогою фазово-контрастної мікроскопії[10].
Просторова варіація показників заломлення в клітині відповідає скупченню розсіювачів різних розмірів, тому будь-яка взаємодія світла з біологічною тканиною характеризується мікроскопічними процесами розсіяння світла [11]
Теорія розсіяння Мі для сферичних частинок будь-якого розміру
Розсіяння світла – це фізичне явище, характерне для світла, що поширюється через середовища з оптичними неоднорідностями, пов'язаними з просторовими коливаннями в показниках заломлення на мікроскопічному рівні[9] .
Коли світло вдаряється в матеріальний об'єкт, електричне поле світла індукує електричний дипольний момент молекули, що коливається з частотою падаючого світла. Тоді кожна молекула діє як осцилююче точкове джерело, що випромінює світло у всіх напрямках. Потужність перевипроміненої (розсіяної) хвилі є ізотропною (однаковою в усіх напрямках) в площині, перпендикулярній напрямку падаючого поля, а її величина пропорційна (де f – частота хвилі; λ – довжина хвилі) [12]. Цей тип розсіювання, відомий як Релеївське розсіювання, генерується від одиничних осцилюючих молекул або молекул, що формують кластери дуже малих часток, чиї лінійні виміри значно менші, аніж довжина хвилі, тому розсіяні ними промені інтерферують лише конструктивно[9].

В 1908 р. німецький фізик Густав Мі запропонував точний розв'язок рівняння Максвелла для однорідних сфер [13]. Запропонована Мі теорія розсіювання світла сферичною частинкою повністю описує електромагнітної плоскої хвилі на сферичних діелектричних об'єктах, будь-якого розміру та для будь-яких довжин хвиль, таким чином дозволяючи передбачити напрямок та величину енергії світла, що поширюється через і довкола сферичної частинки.[12].
Відповідно з теорією розсіяння Мі, напрямок та інтенсивність світла, розсіяного ізотропною, однорідною та сферичною часткою можуть бути обчислені через співвідношення:
- ,

де Is - інтенсивність розсіяного світла, s – коефіцієнт розсіяння частинки; I0 - інтенсивність падаючого світла; λ – довжина хвилі світла; d – точка вимірювання розсіювання світла[9].
Для сферичної частинки, з діаметром рівним чи більшим від довжини світла, характерне розсіяння світла вперед, яке є майже незалежним від довжини хвилі. Таке розсіяння називається «розсіянням Мі» [12].
Теорія розсіяння Мі застосовна до сфер будь-якого розміру, однак є й простіші розв'язки для конкретних випадків, хоч і не застосовні до інших ситуацій. Такою є, напр., теорія Релеївського розсіювання, що є справедливою тільки для малих сферичних часток, з діаметром, меншим ніж 1/10 довжини хвилі світла (d << λ) і яка є обмеженим випадком теорії Мі[9]. Також існує відносно більш простий розв'язок теорії Мі для сферичних часток із значно більшим діаметром, ніж довжина хвилі (d>>λ). В цьому обмеженому випадку використовується класична геометрична оптика (до прикладу, теорія розсіяння Релея-Ганса-Дебая), яка коректно описує поведінку світла на таких частинках в термінах світлових променів[9].
З іншого боку, теорія розсіяння Мі менш коректно передбачає розсіяння світла для неоднорідностей з несферичною формою, випадки кутового розподілу розсіяного світла та багаторазово розсіяне світло[9].
В цілому, на основі співвідношення між розміром частинки та довжиною хвилі світла, процеси розсіяння можна розділити на три різні складові теорії Мі. Це співвідношення визначається, як параметр або розмір Мі[9]:

де k – хвильове число; а – розмір частинки (a = d; де d – діаметр сферичної частинки).
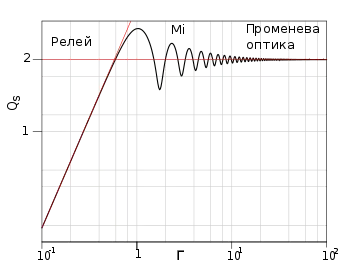
Малюнок подає коефіцієнт ефективності розсіяння, Q (ефективна площа розсіяння), σ, нормована на геометричний поперечний переріз, πr2; де r – радіус сфери) слабко-поглинаючої однорідної сфери , як функцію від параметра Мі[11]. На кривій чітко виділяються три області:
- Релеївське розсіювання: Γ << 1
- Розсіяння Мі: 0,5 < Γ < 10
- Геометрична (променева) оптика: Γ >> 1
Для Релеївського розсіювання, Q швидко зростає із Γ, оскільки σ змінюється пропорційно шостій степені діаметра сфери, d і обернено пропорційно четвертій степені довжини хвилі, λ[12]. Частки в області розсіяння Мі є більш ефективними розсіювачами світла, оскільки світло взаємодіє із частинкою в площі поперечного перерізу, що є більшою від геометричного поперечного перерізу. Це пов'язане із інтерференцією між розсіяними та дифрагованими хвилями, що ілюструють максимальні та мінімальні значення функції. Для більших значень Γ, в області геометричної оптики, ці коливання спадають і Q асимптотично наближається до граничного значення 2. Тут вузьке розсіяння вперед визначається світлом, яке дифрагується довкола частки і світлом, яке прямо падає на неї, що суперечить геометричній оптиці і тому відоме як парадокс екстинкції (ефективна площа розсіяння сфери з великим радіусом в двічі перевищує площу її геометричного поперечного перерізу)[9].
В цілому, частинки з різними розмірами здатні розсіювати світло з різними кутовими розподілами і з різним ступенем ефективності.
Теорія розсіяння Мі і її наближення знайшли широке застосування в обчисленні і моделюванні оптичних властивостей фоторецепторів, зокрема впливу їх складових (ядра, мітохондрій, олійних краплин, структурних неоднорідностей сегментів, мембранних дисків) на ефективність проходження і розсіяння світла. Неоднорідна структура фоторецептора обумовлює його оптичну неоднорідність, що характеризується просторовим коливанням різних показників заломлення, внаслідок чого складові фоторецептора є потенційними елементами розсіяння світла і погіршення зорового зображення [11]. Оскільки діаметр фоторецептора наближається до довжини хвилі видимого світла (~0,5 мкм), то до фоторецепторів застосовні фізичні теорії розсіяння світла на частинках рівних чи більших від довжини хвилі, які дозволяють коректно передбачити проходження, поглинання та розсіяння світла у фоторецепторному хвилеводі.
Розсіяння світла в біологічних тканинах
З оптичної точки зору, біологічна тканина може розглядатись як нерегулярний просторовий розподіл різних показників заломлення, чия величина і масштаб розподілу обумовлені структурою та розміром складових клітин тканини [14].
В біологічній тканині показник заломлення істотно варіюється, від n ≈ 1,33 для води до n ≈ 1,55 для білків та ліпідів[15]. Різними є також показники заломлення деяких складових клітини: клітинного ядра (n ≈ 1,39), мітохондрії (n ≈ 1,41) та цитоплазми (n ≈ 1,38) [16] [17], що робить їх потенційно розсіюючими об'єктами і у випадку фоторецепторів може призводити до розмиття зображення на сітківці (див. нижче «Перевернуте ядро палички»). Так само варіюються геометричні розмірності біологічної тканини: від 1 мкм (субклітинні органели, напр., мітохондрії) до 10-20 мкм (клітина) [8].
Обчислення припускають, що розсіяння в біологічній тканині обумовлені мембранними частинами клітини, зокрема органелами, що містять мембрани, як мітохондрія та ендоплазматичний ретикулум. Причиною такого розсіяння є високий показник заломлення між ліпідами в мембранному бішарі (n ≈ 1,48) та водяній цитоплазмі (n ≈ 1,38) [18].
Велика щільність в тканині розсіюючих об'єктів, чий розмір ближчий чи більший від довжини хвилі світла, призводить до появи множинних «розсіянь Мі», що лише слабо залежать від довжини хвилі. Тому поширення світла в більшості біологічних тканин подібне до поширення світла в довільному середовищі, як густому тумані або хмарах (їх білий колір спричинений розсіянням Мі від краплин води, які мають розмір однаковий чи більший від довжини хвилі, тому розсіяння майже не залежить від довжини хвилі) [19].
Розсіяння світла в оці
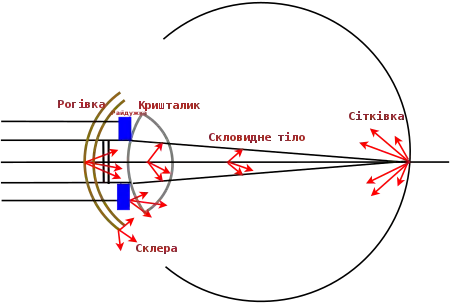
Хоча всі біологічні тканини розсіюють світло, однак око є винятком, оскільки воно повинно забезпечувати фокусування світла і його ефективне поглинання фоторецепторами. Тому деякі елементи здорового, молодого ока (як водяниста волога і скловидне тіло) майже не розсіюють світло. [20]
В ідеальному оці, із цілком прозорими та досконалими оптичними поверхнями не було б жодного розсіяння світла і воно ефективно надходило до фоторецепторів. Однак оптичні елементи біологічного ока хребетних (рогівка, кришталик, скловидне тіло) є потенційними розсіювачами світла, особливо в очах, де функції оптичних елементів порушені внаслідок уражень чи очних хвороб[21]. Тоді як для більшості біологічних тканин характерне «розсіяння Мі», що майже не залежить від довжини хвилі, то для очного середовища характерне розсіяння світла, залежне від довжини хвилі [19].
В людському оці, розсіяння світла призводить до погіршення оптичної якості зображення, обумовлюючи появу відблисків і нечіткого, розмитого зору. Частина світла, яке входить до ока, відхиляється від прямої траєкторії. Але щоб це відхилення впливало на якість зору, розсіяння світла не повинно бути більшим 90°, інакше воно не досягне сітківки і знов вийде з ока внаслідок Релеївського розсіювання (зворотне розсіяння) [20].
Для мінімізації розсіяння світла, що проходить через око до сітківки, кришталик ока є прозорим. Ця прозорість кришталика обумовлена регулярною шестикутною впорядкованістю його складових елементів - видовжених волокон, які мають гладку поверхню і позбавлені клітинного ядра [21]. Строма рогівки, що є прозорим шаром, який складає основну частину рогівки (до 90% її товщини), складається із тонких шарів, паралельних поверхні рогівки. Кожен з цих шарів є 2 мкм в товщину і містить щільно розміщені паралельні один відносно одного колагенові фібрила. Діаметр фібрил ~30 нм, що значно менше ніж довжина хвилі видимого світла. Тож якби ці колагенові фібрила діяли як окремі джерела розсіяння світла, то рогівка була б непрозорою і погіршувала якість зору [22]. Однак ці фібрила розташовані один до одного значно ближче (із міжцентровою відстанню між фібрилам ~50 нм) порівняно з розміром довжини хвилі світла і мають відносно впорядковану, регулярну організацію, тому всі розсіяні хвилі, які не є паралельними падаючому світловому променю, взаємно гасяться через їх деструктивну інтерференцію. На прикладі рогівки кролика було виявлено, що розсіяння світла в цьому оптичному елементі ока сильно залежить від довжини хвилі, як λ-3 або λ -5[20]. Для рогівки ока ця залежність становить λ-4 [23].
Разом з тим, оптичні елементи ока містять ряд неоднорідностей в масштабі порядку довжини хвилі світла (невеликі частинки, шорсткість поверхні, сторонні тіла, коливання щільності), які можуть діяти, як мікроскопічні розсіювачі світла [24]. Подібні мікроскопічні неоднорідності оптики ока здатні зменшувати якість зображення на сітківці і покривати сітківку завісою очного розсіяного світла. Так, частинки в кришталику (т.зв. мультиламелярні тільця, англ. multilamellar bodies), що мають діаметр 1,4 мкм, відповідальні за більшу частину розсіяного вперед світла в оці людини[25][26][27].
Коли прозорість кришталика та рогівки істотно зменшена (напр., внаслідок катаракти чи помутніння рогівки), то ці оптичні елементи стають джерелом як зворотного розсіяння світла, так і розсіяння вперед, внаслідок чого значно зменшується контрастність зображення на сітківці та втрачається гострота зору [24]. Теорія Релеївського розсіювання, разом з іншими наближеними теоріями (як теорія розсіяння Релея-Ганса-Дебая для часток, більших від довжини хвилі світла, поведінка якого тоді описується класичною геометричною оптикою в термінах світлових променів), дозволяють коректно описати розсіяння світла в кришталику ока [28][26].
Іншими джерелами потенційного внутріочного розсіяння світла вперед є райдужна оболонка і склера ока. Хоча їх головною функцією є обмеження надходження світла до ока, вони не цілком непрозорі. Тому, залежно від рівня їх пігментації та щільності, світло може пройти через райдужку та склеру, також призводячи до розсіяння світла [20].
Оскільки окрім розсіяння світла на оптичну якість зображення впливають також аберації фронту хвилі, то розподіл світла в зображенні на сітківці (що описується функцією розсіювання точки), є наслідком складеного ефекту розсіяння світла неоднорідносятми оптики ока та абераціями хвильового фронту[24].
Оптика тканини сітківки
Перевернута сітківка хребетних
В оці хребетних, зображення зовнішнього світу проектуються через оптичну систему ока на світлочутливу сітківку, що простягається уздовж внутрішньої поверхні очного яблука.
Враховуючи мінімізацію розсіяння світла оптикою ока (рогівкою та кришталиком), сітківка хребетних має парадоксальну, перевернуту архітектуру: зображення, спроектовані оптикою ока на сітківку, повинні пройти кілька шарів нейронів, що є безладно орієнтовані та мають нерегулярну форму, перш ніж досягнуть світлочутливих фоторецепторів, які розміщені позаду всіх нейронів сітківки [29] [30].
Хоча, для покращення проходження світла, тонка тканина сітківки є майже прозорою і більшість відростків нейронів сітківки позбавлені мієлінового покриття [31], тим не менше вона складається із кількох нейронних шарів і тому характеризується значним як зворотним розсіянням падаючого світла, так і розсіянням вперед[32].
Структура сітківки хребетних
Сітківка є тонкою (~500 мкм в товщину) прозорою та багатошаровою тканиною, що розміщена позаду ока. Сітківка має високоорганізовану структуру із сімома головними шарами[4].
Три шари містять нейронні клітинні тіла із ядрами (зовнішній ядерний шар, ЗЯШ; внутрішній ядерний шар, ВЯШ; гангліонарний шар, ГШ); два шари складаються із нейронних відростків та синапсів (внутрішній сітчастий шар, ВСШ; зовнішній сітчастий шар, ЗСШ); найбільш глибинний внутрішній шар сітківки містить аксони гангліозних клітин (шар нервових волокон, ШНВ), а найбільш зовнішній шар формується шаром фоторецепторних сегментів (фотосенсорний шар, ФШ), які примикають до пігментного епітелію сітківки (ПЕC) [33].
Розсіяння світла в сітківці
Світло входить до сітківки в ШНВ, а поглинається фоточутливими пігментами фоторецепторів в ФШ, пройшовши всі інші шари. Із ФШ зорова інформація синаптичні передається до біполярних нейронів, які відсилають цю інформацію до ЦНС.
З оптичної точки зору, сітківка складається із множини різноманітних розсіюючих об’єктів. В контексті теорії розсіяння Мі, тканина сітківка розглядається як така, що складається із сферичних часток різного розміру. Параметр Мі, Г, для таких одиничних сфер істотно коливається між ядерними та синаптичними шарами сітківки[11]. Синаптичні структури, чий розмір майже співмірний з довжиною хвилі видимого світла, потенційно можуть призводити до сильного бічного розсіяння. Так само великі клітинні ядра нейронів сітківки є потенційними розсіювачами падаючого світла.
Хоча часто стверджується, що сітківка є майже прозорою, вже той факт, що вона є видимою у водному розчині вказує на розсіяння в тканині сітківки світла внаслідок мікроскопічних фазових варіацій[32]. Крім того, ще в ранніх дослідження оптики сітківки було виявлено, що розсіяння світла в оці, відоме як ентоптичне розсіяння ( ефект «щітки Боема»), на ~40% обумовлене сітківкою, яка тому описувалась як «мутне (оптичне) середовище» (нім. trübes Medium), складене із «світлорозчеплюючих частинок» {lang-de|lichtsplitternden Partikeln } [34][30]. Це узгоджується із пізнішими відкриттями, що із потрапляючого до сітківки видимого світла, тільки ~85% проходить до фоторецепторного шару [35][29].
Найбільше розсіяння світла відбувається в синаптичних шарах сітківки (внутрішньому та зовнішньому сітчастих шарах), де синаптичні елементи мають розмір 500 нм, що наближається до довжини хвилі видимого світла (400-700 нм) і тому призводить до істотного розсіяння падаючого світла[36][37].
Розсіяння падаючого світла в сітківці повинно призводити до появи значного фонового шуму на рівні мозаїки фоторецепторів і, як наслідок, до зменшення співвідношення сигнал/шум (S/N) на рівні зорового сприйняття. Однак зорове сприйняття більшості хребетних має високу роздільну здатність завдяки сітківці, наближаючись до фізичних меж: воно характеризується ефективним контрастним і кольоровим баченням за денного світла та надзвичайно високою чутливістю до світла за нічного, тьмяного освітлення[38]. Тому розсіяння світла в сітківці повинно мінімізуватись оптичними елементами, що допомагають поширенню світла через шари сітківки до місцезнаходження зорових пігментів у фоторецепторі [32].
Вважається, що мінімазація розсіяння світла в сітківці хребетних пов'язана з тим, що вона складається із послідовно впорядкованих біологічних «оптичних волокон», які постають як хвилевідні елементи і компенсують недоліки перевернутої архітектури[32]. Зовнішній та внутрішній сегмент фоторецепторів є одним з прикладів таких хвилевідних елементів, які забезпечують ефективну передачу світла з мінімальним розсіянням до зорового пігменту. Іншими елементами, що мінімізують розсіяння світла в сітківці вважаються гліальні клітини Мюллера та клітинні ядра паличок [39][11].
Оптичні властивості фоторецепторів
Апертура фоторецепторів
Після потрапляння до сітківки, світло спрямовується через шари нейронів до фоторецепторів, в яких воно проходить через внутрішній сегмент (ВС) до зовнішнього сегменту (ЗС), де знаходяться фотопігменти. Оскільки фоторецептори збирають світло на конечній просторовій ділянці (т.зв. вхідній апертурі фоторецептора), вони виконують певного роду просторову інтеграцію падаючих фотонів. Це явище, відоме як ефект апертури, призводить до розмиття зображення на сітківці[40].
Розмір апертури колбочок має важливе значення, оскільки колбочки повинні виявляти різниці в контрастності зображення на сітківці, але разом з тим вони також мають зберігати важливі просторові деталі зображення. Велика апертура має нижчу просторову роздільну здатність, - оскільки збирає всі фотони, які до неї потрапляють, - та усереднює просторові деталі. З іншого боку, велика апертура забезпечує більшу чутливість фоторецептора, оскільки може збирати більше фотонів[41]. Тож хоча наявність конечної апертури фоторецептора може мати негативний вплив на якість зображення на сітківці, апертура також обумовлює скільки світла фоторецептор може поглинути. Чим більше світла поглинається фоторецептором, тим більше інформації про розрізнення передається нейронам сітківки. Це пов'язане з тим, що фотонний шум є пуасонівським процесом, тому S/N збільшується із збільшенням схоплення фотонів. Крім того, у порівнянні із оптикою ока, розмивання зображення на сітківці апертурою фоторецептора є значно меншим[42].
Дослідження з допомогою інтерференційно-контрастного мікроскопа Номарського та психофізіологічні вимірювання апертури показали, що фоторецепторна апертура розміщена на рівні ВС, де показник заломлення паличок та колбочок різниться від позаклітинного простору і фоторецептори оптично відокремленіодин від одного. [43][44] [45]. Тому більшість з того світла, яке входить до ВС колбочок, буде спрямоване в ЗС і діаметр апертури дорівнюватиме діаметру ВС. Відтак, мозаїка колбочкових внутрішніх сегментів, які у центральній ямці щільно розміщені одна до одної, постає також мозаїкою дискретизуючих елементів, які збирають світло та покривають площину зображення на сітківці.
З усіх фоторецепторів, тільки колбочки в центральній ямці мають відносно прямий доступ до зображення на сітківці, оскільки в цьому регіоні внутрішня сітківка та судинна система відсунуті в бік. Діаметр ВС колбочок в макак посередині центральної ямки становить приблизно 2,5 мкм, а з ексцентриситетом в 20 градусів від центральної ямки, діаметр ВС вже складає 8 мкм і ще більше, 11 мкм, на краю скроневої половини сітківки. Діаметр ВС паличок макаки також збільшується від 1,5 мкм одразу після фовеоли до більше 4 мкм біля скроневого краю сітківки, маючи апертуру ~12′′ (12 сек дуги) [46] Оскільки діаметр ВС колбочки на відміну від палички суттєво збільшується з ексцентриситетом, то ефект апертури колбочки також збільшується з ексцентриситетом, тоді як ефект апертури палички залишається майже однаковим. Розмір апертури колбочок в центральній ямці сітківки людини був визначений психофізично і складає приблизно половину анатомічного діаметра колбочки, хоча ймовірно це лише нижча межа її істинного значення [45].
Оптичні властивості ВС колбочок центральної ямки визначають їх здатність до збирання світла, тому збільшення внутрішнього діаметру ВС (при фіксованому міжколбочковому інтервалі) збільшує схоплення ними падаючих фотонів та зменшує невизначеність щодо їх інтенсивності , спричинену фотонним шумом. Потенційно негативним наслідком збільшення діаметру ВС є розмивання зображення внаслідок інтегрування схоплюваного світла апертурою колбочки[47][43]. Втім, відповідно з анатомічними даними, ВС колбочок центральної ямки сітківки людини заповнюють майже весь простір між ними[48] . Це, скоріш за все, свідчить про те, що шкода для якості зображення, спричинена інтегруванням світла апертурою, є незначною за повсякденних візуальних умов [49].
Те, що розмір колбочки в центральній ямці не обмежує якість зображення пов'язане з тим, що апертура колбочки є меншою у порівнянні з функцією розсіювання точки зображення характерною для ока. Тоді як функціональна апертура колбочки в центральній ямці не більша 2,5 мкм в діаметрі чи менша, то функція розсіювання точки ока, навіть при найкращих зорових умовах, становить ~5 мкм в ширину на половинній висоті. Відтак, втрата контрасту, пов'язаного з апертурою колбочки, становить не більше 25% навіть при 60 циклах /градус-1 [49].
Оптична густина фотопігменту
Ізомеризація якомога більшої кількості фотопігменту фотонами, схопленими фоторецепторними апертурами, забезпечує високу зорову чутливість. Однак не все світло, що надходить до фоторецептора, однаково ефективно поглинається фотопігментом.
Здатність середовища поглинати світло характеризується його оптичною густиною, D, що визначається через десятковий логарифм відношення інтенсивності падаючого світла I0 до інтенсивності пройшовшого світла (коефіцієнт пропускання), IT.

Чим вища оптична густина, тим менший коефіцієнт пропускання. [50]
Відповідно до закону Бугера — Ламберта — Бера, оптична густина фотопігменту в фоторецепторі залежить від концентрації фотопігменту в ЗС, c0p, довжини ЗС, l, та коефіцієнту екстинкції фотопігменту (ймовірності поглинання фотону), ε(λ),

де c0 p- концентрація фотопігменту в адаптованому до темряви оці, p – частка не знебарвленого фотопігменту[42]. Фоторецептори поглинають ~50% падаючих фотонів максимальної довжини хвилі світла (λmax)[50].
Осьова оптична густина ЗС фоторецептора є добутком довжини ЗС та питомої густини фотопігменту [51] [52]. Довжина ЗС фоторецепторів, а відтак їх поглинальна здатність, змінюються із збільшення ексцентриситету. Якщо в адаптованому до темряви оці концентрація фотопігменту , c0, є постійною для даного класу фоторецептора , то збільшення довжини ЗС збільшує величину світла, яка може бути накопичене фоторецептором, що призводить до зростання співвідношення сигнал/шум (S/N) [42].
Осьова оптична густина фотопігменту для колбочок, але не для паличок, істотно спадає із ексцентриситетом, як результат морфологічних змін колбочок: периферійні колбочки більш коротші з меншими ЗС, ніж колбочки в центральній ямці. ЗС колбочок є найдовшими у центральній ямці і тому мають тут найбільшу концентрацію фотопігменту на одиницю площі сітківки [53][54]. Довжина ЗС, а відтак і осьова оптична густина фотопігменту, спадає із відстанню від центральної ямки сітківки, коливаючись в діапазоні від 40 до 60 мкм в центральній ямці для L- та M-колбочок, і спадаючи до ~10 мкм на периферії[55].
Зміни в осьовій оптичній густині не впливають на пікову чутливість фотопігменту, однак вони змінюють його спектральну поглинальну здатність, а відтак і спектральну чутливість, що врешті вливає на спектральну світлову ефективність. Чим більшою є оптична густина фотопігменту, тим ширшим є спектр поглинальної здатності, а відтак ширшим є і крива спектральної чутливості. Ширші криві чутливості дозволяють схоплювати більше фотонів, тоді як звуження кривих спектральної чутливості може покращити розрізнення кольорів [56][57] [58]. Це пояснює, чому колбочки на периферії сітківки, які мають низьку оптичну густину у порівнянні із колбочками посередині центральної ямки, характеризуються більш звуженими кривими спектральної чутливості [59].
Наближене значення оптичної густини фотопігменту в даному фоторецепторі може бути отримане із різниць між спектральними чутливостями чи відповідностями кольорів, що вимірюються за нормальної та зменшеної концентрації фотопігменту. Для цього використовують або психофізичні методи (порівняння даних, отриманих коли фотопігмент знебарвлений чи не-знебарвлений або ж оцінки ефекту Стайлса-Кроуфорда), або об'єктивні методи (мікроспектрохімію чи денситометрію сітківки). Отримані психофізичними методами значення середньої пікової оптичної густини для L- та M-колбочок в центральній ямці сітківки людини коливаються від 0,3 до 0,6 (при вимірюванні фотопігменту в стані знебарвлення) та від 0,7 до 1,0 (при аналізі ефекту Стайлса-Кроуфорда) [60][61][62][63][54][64]. Тоді як отримані з допомогою об'єктивних методів значення середньої пікової оптичної густини фотопігменту для L- та M-колбочок коливаються від 0,35 до 0,57[65][66][67][68][59].
Тож, відповідно до психофізичних та об'єктивних методів, осьова оптична густина фотопігменту в ЗС колбочок істотно варіюється між індивідами. Однак в центральній ямці (зокрема в фовеолі) не було виявлено істотної різниці між оптичною густиною M- (0,66) та L-колбочок (0,65), які є винятково ефективними, поглинаючи ~78% фотонів [69].
Визначити оптичну густину фотопігменту S-колбочки значно складніше, оскільки світло, яке його знебарвлює, може призвести до повної втрати чутливості цими колбочками [70]. Опосередковано, на основі порівняння анатомічних властивостей L-, M- та S-колбочок та психофізичних методів, було встановлено, що середня пікова оптична густина фотопігменту S-колбчоок повинна становити ~0,4 для центральної сітківки (з ексцентриситетом 1 градус), та ~30 для периферії (ексцентриситет 13 градусів) [55][71]
Фотопігмент палички, що характеризується рівномірним розподілом поза центром сітківки, має меншу оптичну густину, ніж колбочки, не перевершуючи 0,2 логарифмічних одиниць.[72].
Самоекранування
Вплив оптичної густини фотопігменту на спектральну чутливість фоторецептора пов'язаний з процесом «самоекранування» (англ. «self-screening»), суть якого полягає в тому, що наявність великої кількості молекул фотопігменту в ЗС фоторецептора змінює його загальну спектральну чутливість[73].
Фотони різних довжин хвиль, які проходять по напрямку осі фоторецептора до ЗС, поглинаються неоднорідно: фотони з довжинами хвиль, ближчими до піку чутливості фотопігменту, мають більшу ймовірність бути поглинутими верхніми молекулами фотопігменту, а фотони з іншими довжинами хвиль будуть поглинуті нижче в ЗС. Таким чином, фотопігмент фільтрує падаюче світло, яке проходить через фоторецептор. Внаслідок цього крива спектральної чутливості фоторецептора є ширшою аніж крива чутливості фотопігмента, вимірюваного в розбавленому розчині. Зростання оптичної густини фотопігменту приводить до того, що крива чутливості колбочки стає ширшою. Відтак, до прикладу, дві колбочки, що мають той самий фотопігмент з різними оптичними густинами, матимуть і різні спектральні чутливості[74]. Порівняння вихідних сигналів цих двох колбочок лежить в основі дискримінації кольорів. Крім того, колбочка з вищою оптичною густиною є також більш чутливою по всьому видимому спектру (оскільки більше фотопігменту означає більш фотоізомеризацій).
Ефект Стайлса-Кроуфорда
Зображення на сітківці формується із світла, що надходить з багатьох різних місць в зіниці. Якість цього зображення залежить як від властивостей кришталика та рогівки ока, так і від того, скільки світла, що надійде до кожного місця (точки) в зіниці буде схоплене фоторецепторами. Характерною рисою зору, відмінною від штучних світлочутливих фотоелементів, є чутливість фоторецепторів сітківки до напрямку променя світла[75].
Чутливість фоторецепторів нормального ока є найбільшою по відношенню до світла, що входить до ока біля центру зіниці і спадає майже симетрично, коли точка входу світла зміщується в напрямку до периферії зіниці. Така зміна відносної світлової ефективності як функція від положення на сітківці називається «ефект Стайлса-Кроуфорда І порядку» (ЕСКІ), які вперше її виявили психофізично в 1933 р[76] .
Eксперимент Стайлса і Кроуфорда показав, що коли світло входить до ока в позаосьовій точці зіниці, воно є менш ефективним у генеруванні зорового сприйняття внаслідок того, що він падає під косим кутом на сітківку. При денному світлі, аби генерувати те саме суб'єктивне відчуття яскравості, що й промінь, який входить через центр зіниці, то промінь, який входить через периферію зіниці, повинен мати в 5 - 10 раз більшу енергетичну світність[77]. За 2-3 мм від місця пікової чутливості на сітківці, світло на половину менш ефективне у порівнянні із оптимальним положенням.
Пізніші теоретичні та експериментальні дослідження переконливо показали, що ключовою причиною ЕСКІ є не властивості оптики ока (рогівки, кришталика чи зіниці), а унікальна оптика фоторецепторів, зокрема хвилевідні властивості колбочок, оскільки їх форма та показник заломлення надають їм чутливість до напрямку світла [78][79][80].
Зовнішні сегменти колбочок паралельні і зорієнтовані майже перпендикулярно до зовнішньої оболонки очного яблука, будучи спрямовані до центру зіниці в нормальних очей, таким чином постійно вирівнюючись з найсвітлішою областю зіниці[81]. Існують чисельні свідчення того, що подібне точне вирівнювання колбочок відносно центру зіниці ока пов'язане з тим, що колбочки є фототропіничими та активно орієнтуються до світла, подібно до соняшника в полі [82][83][84].
Фоторецептор, як хвилевід
Одним із методів вивчення сітківки на мікроскопічному рівні в неушкодженому оці є спрямування на сітківку променя світла і відслідковування тієї невеликої частки, яка відбивається назад на зіницю (техніка, відома як «рефлектометрія очного дна»). Тільки ~1% світла, із того що входить до ока, відбивається назад від сітківки, інше нею або розсіюється, або ж поглинається[85]. З того світла, яке поглинається, лише частка схоплюється фотопігментами. До прикладу, із того світла, яке входить до ВС колбочки, лише близько 30% поглинається фотопігментом[86].
Частка світла, що відбивається назад від ока, обумовлена тим, що дещо плоска тканина сітківки діє подібно до слабкого дзеркала, що має значний спрямовуючий компонент. Причиною цієї спрямованості є довга, тонка структура фоторецепторів, що мають вищий показник заломлення аніж оточуюче їх середовище.[87]. Завдяки цій структурі фоторецептори діють, як біологічні оптоволоконні хвилеводи, ефективно скеровуючи світло уздовж всієї їх довжини шляхом повного внутрішнього відбиття і тому можуть бути оптично ізольованими один від іншого, уникаючи розсіяння світла на сусідні клітини і розмиття зображення. [88] [89] [90] [91] [92][93][94].
Хоча нещодавно була запропонована альтернативна від хвилевідної теорія для пояснення фоторецепторної спрямованості, в основі якої є модель дипольної антени, що розглядає кожен фотопігмент, як окремий емітер або приймач випромінювання. [95].
В 1843 р. німецький лікар та фізіолог Ернст Брюкке вперше вказав на те, що показник заломлення колбочок є вищим від оточуючого їх середовища. [96] Враховуючи вищий показник заломлення колбочок та їх майже циліндричну форму, він припустив, що промені світла, які потрапляють до них під відповідним кутом, повинні багаторазово відбиватись в кожному його сегменті (повне внутрішнє відбиття) аж до фотопігмента у зовнішньому сегменті.[97] Брюкке також припустив, що подібне багаторазове відбиття в циліндричній частині фоторецептора унеможливить втечу променів світла через його поздовжню поверхню і розсіяння оптичного зображення в сітківці, таким чином покращуючи гостроту зору.[98] [99] [100].
Хвилевідність світла в фоторецепторі
Оптичний хвилевід має серцевину із високим показником заломлення, яку оточує оболонка із нижчим показником заломлення, тому промінь світла, що входить до хвилеводу поза критичним кутом, не заломлюється, а безперервно відбивається від стінок хвилеводу допоки не досягне іншого кінця. Крім того, для зменшення перехресних перешкод між оптоволоконними елементами, вони мають бути відокремлені (середовищем з нижчим показником заломлення) одне від одного на відстань, що приблизно дорівнює або є більшою ніж довжина хвилі світла[101].
ВС та ЗС фоторецептора відповідає властивостям оптичного хвилеводу, оскільки їх цитоплазма постає як серцевина, а цитоплазматична мембрана та оточуюча рідина фоторецептора, як оболонка[79]. Фоторецептор має високу концентрацію великих молекул (білків, ліпідів, ліпопротеїнів, які забезпечують вищий рівень показника заломлення у порівнянні із оточуючим проміжним (інтерстиціальним) матриксом. Так, у сітківці примат ЗС паличок мають показник заломлення 1,4, а ЗСи колбочок центральної ямки – 1,419, тоді як значення показника заломлення для проміжного матрикса коливається між 1,334 та 1,347[102]. Відстань між фоторецепторами зазвичай складає ~0,51 мкм, що узгоджується з найменшою відстанню для матеріалу з оптично-волоконними властивостями , яка становить 0,5 мкм (1 довжина хвилі в зеленій частині спектра) [78].
Хвилевідна природа фоторецепторів, є більш важливою для колбочок аніж для паличок [103] З ексцентриситетом ~2,5° від середини центральної ямки розташовані колбочки, що характеризуються найбільш вибірковим спрямуванням світла[104].
Хвилевідні моди фоторецептора
Коли світло поширюється через фотоерцептор, чий діаметр наближається до довжини хвилі світла (~0,5 мкм), то оптична поведінка фоторецептора відповідає принципам хвилевідної оптики і тому найкраще описується хвильовою оптикою, а не традиційною геометричною оптикою [105].
Геометрична оптика дозволяє описати як оптичне волокно може схопити та скеровувати промені світла, однак вона не в змозі точно охарактеризувати властивості тих оптоволокон, чий діаметр наближається до довжини хвилі видимого світла, де важливими є ефекти дифракції. Оптичні властивості таких волокон найкраще описуються хвильовою оптикою, яка замінює промені у хвилеводі на електромагнітні поля. Коли хвилі світла поширюються в хвилеводі, то інтерферують, що призводить до формування всередині та довкола хвилеводу стабільних форм коливань світла – мод, кожна з яких пов'язана із характерним тривимірним розподілом густини енергії в хвилеводі[101]. Такі моди є єдиним способом, яким світло поширюється через хвилевід і вони можуть бути згруповані в кілька порядків, пов'язаних з числом модальних поверхонь, на яких електромагнітні поля прямують до нуля: мода першого порядку (основна), другого, третього, четвертого і т.д. Порядок моди, яка поширюється в хвилеводі, залежить від його діаметру: менші моди (нижчого порядку) поширюються у тонших світловодах, а збільшення діаметру призводить до поширення все більших порядків мод. Однак основна мода поширюється завжди, незалежно від діаметра хвилеводу[2].

В кінцевій точці хвилеводу, волокно постає як антена, випромінюючи енергію світла. Спостерігаючи такий кінець хвилеводу під мікроскопом можна побачити більш менш складні форми світла, які через передані моди приблизно відтворюють поперечний перетин волокна. Одним з перших, хто побачив хвилевідні моди в фоторецепторах був данський фізіолог Адольф Ганновер в 1843, хоча і вважав їх лише витонченими клітинними структурами [106]. Тільки з відкриттям ефекту Стайлса-Кроуфорда І відродилась цікавість до напрямкових властивостей фоторецепторів та намагання з їх допомогою пояснити ефект. На початку 1960-х рр. вперше були виявлені хвилевідні модальні структури у фоторецепторах, яких було знайдено 12 [88][90] [91]
Оптичні властивості ідеального хвилеводу, що має поперечний діаметр d (у випадку фоторецептора – ефективна апертура) та показник заломлення n1, і оточений середовищем з показником заломлення n2 (<n1), цілком описуються безрозмірним «хвилевідним параметром», V:

де λ – довжина хвилі світла. Для даного хвилеводу V змінюється із зміною довжини хвилі. За винятком моди найнижчого порядку, кожна з мод хвилеводу пов'язана з порогом, Vc, нижче якого світло не може поширюватись в такій моді волокном нескінченно.
Для фоторецепторів, не враховуючи основні моди, які завжди поширюються, для інших допустимих мод V знаходиться в діапазоні між 2,4 та ~5,5[91]. Для міоїда та ЗС колбочок центральної ямки хвилевідний параметр V = 3,38 та 2,58 відповідно [107]. Відтак, тільки моди HE11 (2 моди), TE01 (1 мода), TM01 (1 мода) та HE21 (2 моди) мають нижчі граничні значення і тому можуть поширюватись через міоїд, еліпсоїд та ЗС фоторецептора.
Важливою властивістю мод є те, що не вся їх енергія поширюється всередині світловоду. Частина енергії моди поширюється поза хвилеводом і чим більший порядок моди, тим більшою є ця частка. Це істотно впливає на просторову роздільну здатність зору: чим більше світла поширюється поза даним фоторецептором, тим більше світла поглинається сусідніми з ним фоторецепторами. Такі фоторецептори стають оптично поєднаними, що погіршує роздільну здатність зору. Тільки енергія мод в межах фоторецептора взаємодіє з фотопігментом. [108] [109]
Функції хвилевідності
Хвилевідна природа уможливлює високе поглинання фоторецептором світла при відносно малих затратах фотопігменту, оскільки поглинання фотонів залежить головним чином від довжини фоторецептора і значно менше від його діаметра та форми. [78] Найменший можливий діаметр фоторецептора визначається ефектом оптичного хвилеводу. Нижче критичного діаметру в середину хвилеводу потрапляє менше світла, що робить його менш ефективним[110].
Хвилевідні властивості колбочок обумовлюють залежність ефективності схоплення фотонів сітківкою від точки входу світла в зіницю (ефект Стайлса-Кроуфорда І типу). Щойно світло входить до ВС колбочки, то внутрішні відбиття від стінок фоторецептора спрямовують світло вздовж його довгої осі до ЗС, де міститься фотопігмент. Одним із наслідків цього є більш ефективне схоплення та використання апертурами фотонів, оскільки вони можуть бути зібрані вздовж більшого діапазону положень та кутів променя світла. Світло, що прямо входить у ВС найкраще схоплюється фоторецептором та спрямовується до ЗС, тоді як світло, яке надходить до ВС під більшими кутами, хоча і може бути схоплене, але матиме меншу ефективність у проходженні до ЗС [108][2].
Хвилевідність також є ефективною для світла, яке через фоторецептор відбивається назад, оскільки оптичні системи є зворотними. Колбочки мають два головні відбиваючі компоненти: 1) анатомічне з’єднання між ВС та ЗС, та 2) з’єднання між ЗС та пігментним епітелієм сітківки. Відтак, іншим наслідком хвилевідної природи фоторецептора є те, що внутрішні відбиття світла здатні знову спрямувати вздовж прямого шляху входження ту його частину, яка вдаряється у ці з’єднання та може розсіятись.[87]
Хвилевідні властивості фоторецепторів є результатом еволюції їх тонких, видовжених структур, які самі постали в процесі оптимізації витрат фотопігменту та квантового поглинання[110].
Дискретизація зображення фоторецепторами
Оброблення зорової інформації розпочинається вже на рівні оптики ока, яка складається із трьох головних компонент: рогівки, зіниці та кришталика. Ці оптичні компоненти формують чітке зображення на рівні фоторецепторів, які дискретизують неперервне зображення на сітківці і перекодовують його в електричні сигнали.
Правило Гельмгольца
Щільність розміщення колбочок в центральній ямці є важливою для забезпечення високої роздільної здатності зору. Найменша кутова відстань між сусідніми об'єктами, яку може розрізнити зір, називається «мінімально-розрізнюваною гостротою» зору [111]. Мінімально-розрізнювана гострота зору оцінюється через визначення найчіткіших чорних та білих смуг, які можуть бути розрізнені спостерігачем. За ідеальних умов (при високій контрастності та яскравості) людина з відмінним зором може розрізнити ґратки змінюваних чорних та білих смуг, коли один цикл утворює кут в ~1 мінуту дуги (0,017 кутових градусів). Така мінімально-розрізнювана гострота є одним з головних обмежень просторового зору, будучи найчіткішою деталлю високої контрастності, що може бути розрізнена людським оком. Для зору, що спирається на центральну ямку сітківки, однією з головних причин такого обмеження є інтервал між колбочками.
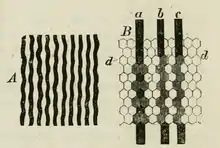
Зовнішній світ, доступний зору, представлений як безперервна варіація в інтенсивності світла. Однак візуальна система, дивлячись на розподіл світла через одиничні фоторецептори, формує із безперервної варіації дискретні «вибірки». Тому початком процесу бачення є трансформація (дискретизація) щільною мозаїкою фоторецепторів безперервного зображення на сітківці в дискретні вибірки. Чим більш щільно розміщені фоторецептори, тим краще зорова система може відображати розподіл світла[112].
Те, що зір починається із просторової дискретизації зображення на сітківці дискретними фоторецепторами, вперше відзначив в 1850-х рр. німецький анатом Генріх Мюллер, коли виявив, що палички та колбочки є світлочутливими елементами сітківки. Однак сам Мюллер спочатку вважав, що для того, щоб зір сприймав дві точки світла як окремі , достатньо щоб ці точки стимулював дві окремі колбочки[113]. Проте, як відзначив Карл Бергман (фізіолог) (1858), в такому випадку зір не зможе відрізнити безперервну лінію від ряду точок[114].
Бергман, а згодом учень Г.Мюллера, Герман фон Гельмгольц (1860) висунули іншу гіпотезу (сьогодні відому як «правило Гельмгольца»): для розрізнення двох точок, потрібно щоб один не стимульований фоторецептор був розміщений між двома стимульованими фоторецепторами[115][116]. Хоча це правило було сформульоване у контексті розрізнення двох точок, воно також застосовне у випадку розрізнення одиничних смуг синусоїдальних ґраток. Для випадку таких ґраток із чорних та білих смуг, правило Гельмгольца постулює необхідність існування двох точок вибірки на один цикл ґратки, тоді чорні та світлі смуги можуть окремо стимулювати фоторецептори.
Межа Найквіста
Зображення ґратки, яке відповідно до правила Гельмгольца узгоджується із дискретизуючою мозаїкою, знаходиться на частоті зрізу цієї мозаїки, fc, оскільки є найвищою просторовою частотою, яку система може розрізнити та надіслати до вихідних елементів. Ґратка, що має нижчу просторову частоту аніж частота зрізу, хоч і передискретизується, але уможливлює генерування множини вихідних значень, що без спотворень визначають вхідний стимул та дозволяють його точно реконструювати, fc/2. Тільки ґратка, що має частоту, вищу від частоти зрізу (3fc ), субдескритизується системою та призводить до втрати інформації дискретизуючим елементами. В цьому випадку вхідний стимул не може бути реконструйований і передається як сигнал із нижчою частотою. Таке спотворення інформації відоме як аліасинг [40].
Гіпотеза Бергмана та Гельмгольца була чітко формалізована через півстоліття у відомій в теорії інформації «теоремі дискретизації» (або теорема відліків) у відповідності з якою, для відновлення одновимірного сигналу з дискретної послідовності n рівновіддалених вибірок , частота дискретизації повинна бути рівною або у два рази більшою від максимальної частоти сигналу, fc:fc=n/2[117][118]. Інакше кажучи, теорема постулює, що в результаті дискретизації безперервного сигналу ніяка інформація не буде втрачена, за умови якщо процес дискретизації безпомилковий і найвища частота в сигналі становить не більше половини частоти дискретизації. Частота зрізу (половина частоти дискретизації) називається межею Найквіста[119].
У застосуванні до фоторецепторів теорема дискретизації припускає, що максимальна просторова частота, що може бути реконструйована (дискретизована) без спотворень, повинна вдвічі перевищувати інтервал між фоторецепторами. Тому для будь-якого даного інтервалу між фоторецепторами існує верхня межа (межа Найквіста) і будь-які просторові частоти в безперервному зображенні, що перевершують цю межу, будуть спотворені, як такі що мають нижчу просторову частоту (аліасинг).
Межа Найквіста для фоторецепторів обчислюється із середнього інтервалу між фоторецепторами, хоча однозначно визначеною вона є лише для випадку ідеально впорядкованих мозаїк, тоді як навіть мозаїка колбочок в центральній ямці далека від такої регулярності.[47] Для ока людини межа дискретизації Найквіста становить приблизно 60 циклів/градус, стільки ж скільки й вища просторова частота зрізу оптики ока. Людське око не в змозі виявити частоти вище ніж 60 циклів/градус, оскільки частота дискретизації колбочок в центральній ямці становить ~120 циклів/градус [120][121].
Тільки окремі види птахів мають гостріший зір, аніж примати. Так, орлан білоголовий має гостроту вище 120 циклів/градус, що досягається не шляхом більш щільного пакування фоторецепторів, а шляхом більш глибшої центральної ямки, завдяки чому колбочки сітківки орлана схоплюють вищі просторові частоти, аніж колбочки людської сітківки.
Аліасинг
Кодування фоторецепторами зображення у формі дискретних вибірок має два важливі наслідки для зору. Перший полягає в тому, що зорова система стикається з проблемою аліасинга: при реконструкції безперервного оптичного зображення із значень дискретних вибірок, розбіжність між смугою пропускання зображення та частотою дискретизації приводить до появи просторових ілюзій, коли високі просторові частоти (вимірюється в циклах на кутовий градус) первинного зображення при його реконструкції постають як низькі просторові частоти [122]. Іншим наслідком є те, що інтервал між одиничними вибірками, що відповідає інтервалу між фоторецепторами, накладає обмеження на просторову роздільну здатність зору. Причиною аліасинга, - який по суті є формуванням низькочастотного муарового візерунку, - є субдескритизація зображення на сітківці, що має місце тоді, коли частота дискретизації регулярної мозаїки точок в два рази менша від найвищої просторової частоти, що відображена на цій мозаїці.
Застосування теореми дискретизації до мозаїки колбочок демонструє, що максимальна просторова частота, яка може бути відтворена без спотворень (аліасинга), рівна половині частоти дискретизації мозаїки колбочок. Однак хоча теорема дискретизації часто застосовується до мозаїки колбочок, ця мозаїка не є ні регулярною, ні одновимірною, будучи двовимірним нерегулярним дискретизуючим масивом. Хоча для цього випадку також існує багатовимірна теорема дискретизації [123]. Але теорема дискретизації для одновимірного випадку підходить для колбочок в центральній ямці, які мають достатньо регулярну мозаїку, яка здатна генерувати муарові візерунки, схожі на смуги зебри. Поза центральною ямкою аліасинг постає як двовимірний просторовий шум [112]
У випадку, коли мозаїка колбочок дискретизує два синусоїдальні стимули, зорова система не в змозі визначити, чи отримані дискретні вибірки із нижчих частот, що були дискретизовані достатньою кількістю фоторецепторів, чи ці вибірки із вищих частот, що були субдескритизовані. У випадку субдескритизації зображення на сітківці мозаїкою фоторецепторів, просторові частоти вище межі Найквіста хибно тлумачаться зоровою системою як низькочастотні, породжуючи «змішані ефекти». Зорова система не в змозі відрізнити такі змішані ефекти від природних низьких просторових частот і тому спотворює дискретизовану репрезентацію зображення на сітківці, інтерпретуючи субдескритизовані зображення наче вони чіткі[124].
Спектр просторових частот зображення на сітківці визначається просторово-частотним спектром візуальної сцени та модуляційною передавальною функцією. Хоча спектри просторових частот природних візуальних сцен варіюються, в цілому вони мають амплітуди, які спадають обернено пропорційно просторовій частоті, що саме по собі допомагає зоровій системі уникати змішаних ефектів (аліасинга) [125]. Аліасинг виникає тільки у випадку, коли модуляційна передавальна функція перевищує межу Найквіста.
Оскільки аліасинг не може одразу бути виправлений процесами що мають місце після дискретизації, він накладає фундаментальні обмеження на будову систем передачі зображення. Загальноприйнятим вирішенням цієї проблеми в інженерії є недопущення надходження просторових частот вище межі Найквіста до дискретизуючого масиву. Це здійснюється через попередню фільтрацію зображення, тобто обмеження смуги частот зображення до початку дискретизації, коли частоти, що призведуть до аліасинга, відфільтровуються оптичною передавальною функцією камери [126].
На відміну від інженерних систем, зорова система людини не вирішує проблему аліасинга шляхом обмеження смуги частот зображення, а подекуди навіть його допускає. Більшій частині сітківки притаманна значна невідповідність між границями смуги пропускання зображення на сітківці, накладеними оптикою ока, та частотами Найквіста, що випливає із загальної кількості фоторецепторів, доступних для дискретизації цього зображення[127]. Винятком є середина центральної ямки (фовеола), де середня міжрецепторна відстань (від центру до центру) складає (120)-1 градусів кута поля зору [128] . Позаяк в центральній ямці межа Найквіста становить ~60±10 циклів/градус зорового кута, що наближається до найдетальнішої структури, яка може бути сприйнята оптичним апаратом ока за оптимальних умов, то зір людини що спирається на колбочки центральної ямки, добре захищений від аліасинга оптичними властивостями ока, які відфільтровують просторові частоти, що перевершують межу Найквіста мозаїки колбочок центральної ямки[129][130]. Внаслідок такого фільтрування людина не в змозі візуально сприймати властивості зовнішнього світу (на рівні атомів та молекул), виражені в просторових частотах вище 60 циклів/градус.
Фовеола є єдиною областю сітківки ока людини, якій притаманна чітка підігнаність між межею Найквіста фоторецепторної мозаїки та шириною смуги пропускання зображень, які можуть бути сформовані на цій мозаїці при нормальнім зорові. Гельмгольц вперше висловив думку, що таке співвідношення між оптикою ока та мозаїкою фотоерцепторів свідчить про їх підігнаність (узгодженість) одне до одного [116]. Хоча це в першому наближенні справедливо для мозаїки колбочок в фовеолі, поза цією областю оптика ока істотно перевершує роздільну здатність периферійної мозаїки колбчок. Із збільшенням ексцентриситету від центральної ямки, ширина смуга пропускання оптики ока спадає повільніше, тоді як міжцентровий інтервал колбочок швидко зростає, внаслідок чого зображення на сітківці субдескритизовані й периферійний зір повинен сприймати аліасинг. [131] . Однак не зважаючи на невідповідність між шириною смуги пропускання зображення поза центральною ямкою та щільністю фоторецепторів, зорова система здатна запобігати аліасингу.
Однією з причин, чому периферійний аліасинг не є відчутним при повсякденному функціонуванні зору, є різна оптична якість зору поміж різних індивідів. До прикладу, такі аберації периферійної оптики, як астигматизм ока з косими осями значно зменшують силу саме тих просторових частот, що можуть викликати змішані ефекти . Крім того, поєднання ряду інших факторів (напр., розфокусування внаслідок затримки акомодації [133]; незначна присутність в природних сценах високих просторових частот [134]; поперечна хронічна аберація [135]; просторова нерегулярність периферійної мозаїки колбочок через присутність паличок [127] [136]) також зменшують вплив периферійного аліасинга на нормальне функціонування зору.
Крім того, еволюційні механізми повинні були спрямовувати частоту зрізу оптики ока до частот, вищих від межі Найквіста, оскільки покращення контрастності зображення при просторових частотах нижче межі Найквіста більш ніж компенсує згубні ефекти будь-якого аліасинга просторових частот вище цієї межі [137]. Тому однією з переваг субдескритизації мозаїкою колбочок зашумлених зображень на сітківці може бути підвищення різкості зображення на високих просторових частотах. Також зорова система може використовувати з’єднання між колбочками через щілинні контакти для покращення чутливості до нижчих просторових частот без істотного зниження візуальної роздільної здатності чи збільшення ефектів аліасинга[138].
Тож, відповідність між межею Найквіста і найбільшою просторовою частотою, яка може бути виявлена зором, не є нездоланним бар'єром для роздільної здатності людського ока.[139]. Сам по собі процес дискретизації не має впливу на контрастність зображення. Зменшуючи розмірність зображення, дискретизація призводить до появи неоднозначності лише тому, що сигнал між двома точками вибірки невідомий. Тому функцією пострецепторних нейронів є відтворення та інтерпретація двозначного зображення.
Найбільш переконливо на користь того, що межа Найквіста не є нездоланною для ока, свідчить оригінальний експеримент, в якому спостерігачам показувались високочастотні синусоїдальні ґратки, використовуючи лазерну інтерферометрію щоб уникнути зменшення контрастності, що має місце коли стимул проходить через оптику ока[121]. Було виявлено, що спостерігачі могли надійно розрізнити ґратки з частотою до 150 циклів/градус, що в 2,5 перевершує межу Найквіста в позафовеальній сітківці[140]. І хоча дискримінація просторових частот стає надзвичайно поганою для частот, що перевершують межу Найквіста, в цих просторових частотах зберігається незначна інформація про орієнтацію ґраток [112].
Джерела
- Miller W.H. (1974). Receptor-optic wave-guide effects.. Investigative Ophthalmology & Visual Science 13 (8): 556–9. PMID 4841863.
- Snyder A.W (1975). Photoreceptor Optics — Theoretical Principles. У Snyder A.W. & Menzel R. Photoreceptor Optics. Springer. с. 38–55. ISBN 978-3-642-80934-7. doi:10.1007/978-3-642-80934-7_3.
- Snyder A.W and Menzel R. (1975). Introduction to Photoreceptor Optics — An Overview. У Snyder A.W. & Menzel R. Photoreceptor Optics. Springer. с. 1–13. ISBN 978-3-642-80934-7. doi:10.1007/978-3-642-80934-7_1.
- Rodieck, Robert W. (1998). The First Steps in Seeing. Sinauer Associates. с. 562. ISBN 978-0-87893-757-8.
- Burns S.A., Wu S., Delori F., and Elsner A.E. (1995). Direct measurement of human-cone-photoreceptor alignment. Journal of the Optical Society of America A 12 (10): 2329. ISSN 1084-7529. doi:10.1364/JOSAA.12.002329.
- Vohnsen B. (17 лютого 2017). The retina and the Stiles–Crawford effects. У Artal P. Handbook of Visual Optics, Volume One: Fundamentals and Eye Optics. CRC Press. с. 257–276. ISBN 978-1-315-35572-6.
- Lakshminarayanan V. and Enoch J M. (4 December 2009). Biological Waveguides. У Bass M. et al. Handbook of Optics. Volume III. Vision and Vision Optics (вид. 3). McGraw Hill Professional. с. 8.1–8.29. ISBN 978-0-07-162928-7.
- Tuchin V. (2015). Tissue Optics: Light Scattering Methods and Instruments for Medical Diagnosis (вид. 3). SPIE. с. 988. ISBN 978-1-62841-516-2.
- van de Hulst, H. C. (1957). Light Scattering by Small Particles. New York: Dover Publications. с. 470. ISBN 978-0-486-64228-4.
- Zernike F. (1955). How I Discovered Phase Contrast. Science 121 (3141): 345–349. ISSN 0036-8075. doi:10.1126/science.121.3141.345.
- Agte, Silke (2013). [id=10870 Light guidance in Müller cells of the vertebrate retina. PhD thesis]. Universität Leipzig. с. 157.
- Stratton J.A. (22 січня 2007). Electromagnetic Theory. John Wiley & Sons. с. 615. ISBN 978-0-470-13153-4.
- Mie G. (1908). Beiträge zur Optik trüber Medien, speziell kolloidaler Metallösungen. Annalen der Physik 330 (3): 377–445. ISSN 00033804. doi:10.1002/andp.19083300302.
- Drezek R., Dunn A., and Richards-Kortum R. (1999). Light scattering from cells: finite-difference time-domain simulations and goniometric measurements. Applied Optics 38 (16): 3651. ISSN 0003-6935. doi:10.1364/AO.38.003651.
- Bolin F.P., Preuss L.E., Taylor R.C., and Ference R.J. (1989). Refractive index of some mammalian tissues using a fiber optic cladding method. Applied Optics 28 (12): 2297. ISSN 0003-6935. doi:10.1364/AO.28.002297.
- Brunsting A. and Mullaney P.F. (1974). Differential Light Scattering from Spherical Mammalian Cells. Biophysical Journal 14 (6): 439–453. ISSN 00063495. doi:10.1016/S0006-3495(74)85925-4.
- Liu, H., B. Beauvoit, M. Kimura, and B. Chance (1996). Dependence of tissue optical properties on solute-induced changes in refractive index and osmolarity. Journal of Biomedical Optics 1 (2): 200. ISSN 10833668. doi:10.1117/12.231370.
- Beuthan J., Minet O., Helfmann J., Herrig M., and Müller G. (1996). The spatial variation of the refractive index in biological cells. Physics in Medicine and Biology 41 (3): 369–382. ISSN 0031-9155. doi:10.1088/0031-9155/41/3/002.
- Svaasand L.O. (2003). Lasers for biomedical applications. У Driggers R.G. Encyclopedia of Optical Engineering: Las-Pho. CRC Press. с. 1035–1045. ISBN 978-0-8247-4251-5.
- van den Berg, Thomas J.T.P. (17 лютого 2017). Scattering, straylight, and glare. У Artal P. Handbook of Visual Optics, Volume One: Fundamentals and Eye Optics. CRC Press. с. 349–362. ISBN 978-1-315-35572-6.
- Atchison David A. and Smith George (29th February 2000). Optics of the Human Eye. Butterworth-Heinemann. с. 288. ISBN 978-0-7506-3775-6.
- Maurice D.M. (1957). The structure and transparency of the cornea. The Journal of Physiology 136 (2): 263–286. ISSN 00223751. doi:10.1113/jphysiol.1957.sp005758.
- Van Den Berg, T.J.T.P. and Tan, K.E.W.P. (1994). Light transmittance of the human cornea from 320 to 700 nm for different ages. Vision Research 34 (11): 1453–1456. ISSN 00426989. doi:10.1016/0042-6989(94)90146-5.
- Piñero D.P., Ortiz D., and Alio J.L. (2010). Ocular Scattering. Optometry and Vision Science 87 (9): E682–E696. ISSN 1040-5488. doi:10.1097/OPX.0b013e3181e87da6.
- van den Berg TJ (1997). Light scattering by donor lenses as a function of depth and wavelength.. Investigative Ophthalmology & Visual Science 38 (7): 1321–32. PMID 9191595.
- van den Berg T.J.T.P and Spekreijse H. (1999). Light scattering model for donor lenses as a function of depth. Vision Research 39 (8): 1437–1445. ISSN 00426989. doi:10.1016/S0042-6989(98)00220-X.
- Vos J.J. (2003). On the cause of disability glare and its dependence on glare angle, age and ocular pigmentation. Clinical and Experimental Optometry 86 (6): 363–370. ISSN 08164622. doi:10.1111/j.1444-0938.2003.tb03080.x.
- Costello M.J., Johnsen S., Gilliland K.O., Freel C.D., and Fowler W.C. (2007). Predicted Light Scattering from Particles Observed in Human Age-Related Nuclear Cataracts Using Mie Scattering Theory. Investigative Opthalmology & Visual Science 48 (1): 303. ISSN 1552-5783. doi:10.1167/iovs.06-0480.
- Hammer M., Roggan A., Schweitzer D., and Muller G. (1995). Optical properties of ocular fundus tissues-an in vitro study using the double-integrating-sphere technique and inverse Monte Carlo simulation. Physics in Medicine and Biology 40 (6): 963–978. ISSN 0031-9155. doi:10.1088/0031-9155/40/6/001.
- Vos J. J. and Bouman M. A. (1964). Contribution of the Retina to Entoptic Scatter. Journal of the Optical Society of America 54 (1): 95. ISSN 0030-3941. doi:10.1364/JOSA.54.000095.
- Enoch J.M. and Glisman L.E. (1966). Physical and Optical Changes in Excised Retinal Tissue: Resolution of Retinal Receptors as a Fiber Optic Bundle. Investigative Ophthalmology & Visual Science 5: 208–221.
- Reichenbach A., Agte S., Francke M., and Franze K. (2014). How light traverses the inverted vertebrate retina. e-Neuroforum 5 (4): 93–100. ISSN 1868-856X. doi:10.1007/s13295-014-0054-8.
- Masland Richard H. (2012). The Neuronal Organization of the Retina. Neuron 76 (2): 266–280. ISSN 08966273. doi:10.1016/j.neuron.2012.10.002.
- Boehm G. (1940). Über ein neues entoptisches Phänomen im polarisierten Licht beschrieben; »Periphere« Polarisationsbüschel. Acta Ophthalmologica 18 (2): 143–169. ISSN 1755375X. doi:10.1111/j.1755-3768.1940.tb07412.x.
- Boucher F., Leblanc R.M., Savage S., and Beaulieu B. (1986). Depth-resolved chromophore analysis of bovine retina and pigment epithelium by photoacoustic spectroscopy. Applied Optics 25 (4): 515. ISSN 0003-6935. doi:10.1364/AO.25.000515.
- Martins-Ferreira H and de Castro GO (1966). Light-scattering changes accompanying spreading depression in isolated retina.. Journal of Neurophysiology 29 (4): 715–26. PMID 5966432.
- Knighton RW, Jacobson SG, and Kemp CM (1989). The spectral reflectance of the nerve fiber layer of the macaque retina.. Investigative Ophthalmology & Visual Science 30 (11): 2392–402. PMID 2807795.
- Land M.F. and Nilsson D.-E. (1 March 2012). Animal Eyes. Oxford Animal Biology Series (вид. 2). Oxford University Press. с. 271. ISBN 978-0-19-958114-6.
- Reichenbach A., Savvinov A., Wurm A., Grosche J., Guck J., Franze K., Skatchkov S.N., Agte S., and Junek S. (2012). Live Cells as Optical Fibers in the Vertebrate Retina. У Yasin, Moh. Selected Topics on Optical Fiber Technology. INTECH Open Access Publisher. с. 247–270. ISBN 978-953-51-0091-1. doi:10.5772/26945.
- Hughes, A. (18 грудня 1986). The schematic eye comes of age. У Pettigrew, J., Sanderson, K.J. and Levick, W.R. Visual Neuroscience. Cambridge University Press. с. 60–89. ISBN 978-0-521-25829-6.
- Valberg, Arne (3 лютого 2007). Light Vision Color. John Wiley & Sons. с. 474. ISBN 978-0-470-01212-3.
- Geisler W.S. and Banks M.S. (4 December 2009). Visual performance. У Bass M. et al. Handbook of Optics. Volume III. Vision and Vision Optics (вид. 3). McGraw Hill Professional. с. 2.1–2.60. ISBN 978-0-07-162928-7.
- Miller William H. and Bernard Gary D. (1983). Averaging over the foveal receptor aperture curtails aliasing. Vision Research 23 (12): 1365–1369. ISSN 00426989. doi:10.1016/0042-6989(83)90147-5.
- Chen Bing, Makous Walter and Williams David R. (1993). Serial spatial filters in vision. Vision Research 33 (3): 413–427. ISSN 00426989. doi:10.1016/0042-6989(93)90095-E.
- Macleod Donald I.A., Williams David R. and Makous Walter (1992). A visual nonlinearity fed by single cones. Vision Research 32 (2): 347–363. ISSN 00426989. doi:10.1016/0042-6989(92)90144-8.
- Packer Orin, Hendrickson Anita E. and Curcio Christine A. (1989). Photoreceptor topography of the retina in the adult pigtail macaque (Macaca nemestrina). The Journal of Comparative Neurology 288 (1): 165–183. ISSN 0021-9967. doi:10.1002/cne.902880113.
- Snyder A.W. and Miller W.H. (1977). Photoreceptor diameter and spacing for highest resolving power. Journal of the Optical Society of America 67 (5): 696. ISSN 0030-3941. doi:10.1364/JOSA.67.000696.
- Curcio C., Sloan K., Packer O., Hendrickson A. and Kalina R. (1987). Distribution of cones in human and monkey retina: individual variability and radial asymmetry. Science 236 (4801): 579–582. ISSN 0036-8075. doi:10.1126/science.3576186.
- Williams, D.R. (1986). Seeing through the photoreceptor mosaic. Trends in Neurosciences 9: 193–198. ISSN 01662236. doi:10.1016/0166-2236(86)90058-5.
- Hsia, Y. (1965). Photochemistry of vision. У Graham, C.H. Vision and visual perception. New York: Wiley. с. 132–153.
- Baylor D.A., Nunn B.J. and Schnapf L. (1984). The photocurrent, noise and spectral sensitivity of rods of the monkey Macaca fascicularis.. The Journal of Physiology 357 (1): 575–607. ISSN 00223751. doi:10.1113/jphysiol.1984.sp015518.
- Fain G.L., Lamb T.D., Matthews H.R. and Murphy R.L. (1989). Cytoplasmic calcium as the messenger for light adaptation in salamander rods.. The Journal of Physiology 416 (1): 215–243. ISSN 00223751. doi:10.1113/jphysiol.1989.sp017757.
- Marcos S., Tornow R.-P., Elsner A.E., and Navarro R. (1997). Foveal cone spacing and cone photopigment density difference: Objective measurements in the same subjects. Vision Research 37 (14): 1909–1915. ISSN 00426989. doi:10.1016/S0042-6989(96)00295-7.
- Elsner A. E. and Burns S. A. (1993). Mapping cone photopigment optical density. Journal of the Optical Society of America A 10 (1): 52. ISSN 1084-7529. doi:10.1364/JOSAA.10.000052.
- Stockman A., Sharpe L.T. and Fach C. (1999). The spectral sensitivity of the human short-wavelength sensitive cones derived from thresholds and color matches. Vision Research 39 (17): 2901–2927. ISSN 00426989. doi:10.1016/S0042-6989(98)00225-9.
- He J.C. and Shevell S.K. (1995). Variation in color matching and discrimination among deuteranomalous trichromats: Theoretical implications of small differences in photopigments. Vision Research 35 (18): 2579–2588. ISSN 00426989. doi:10.1016/0042-6989(95)00007-M.
- Neitz J., Neitz M., He J.C., and Shevell S. K. (1999). Trichromatic color vision with only two spectrally distinct photopigments. Nature Neuroscience 2 (10): 884–888. ISSN 10976256. doi:10.1038/13185.
- Thomas P.B.M., Formankiewicz M.A. and Mollon J.D. (2011). The effect of photopigment optical density on the color vision of the anomalous trichromat. Vision Research 51 (20): 2224–2233. ISSN 00426989. doi:10.1016/j.visres.2011.08.016.
- Berendschot T.T., van de Kraats J., and van Norren D (1996). Foveal cone mosaic and visual pigment density in dichromats.. The Journal of Physiology 492 (1): 307–314. ISSN 00223751. doi:10.1113/jphysiol.1996.sp021310.
- Alpern M. (1979). Lack of uniformity in colour matching.. The Journal of physiology 288: 85–105. PMC 1281416. PMID 313984. doi:10.1113/jphysiol.1979.sp012685.
- Wyszecki G. and Stiles W.S. (1980). High-level trichromatic color matching and the pigment-bleaching hypothesis. Vision Research 20 (1): 23–37. ISSN 00426989. doi:10.1016/0042-6989(80)90138-8.
- Smith VC and Pokorny J (1973). Psychophysical estimates of optical density in human cones.. Vision Research 13 (6): 1199–202. PMID 4713928.
- Miller S. S. (1972). Psychophysical estimates of visual pigment densities in red-green dichromats. The Journal of Physiology 223 (1): 89–107. ISSN 00223751. doi:10.1113/jphysiol.1972.sp009836.
- Walraven P. L. and Bouman M. A. (1960). Relation between Directional Sensitivity and Spectral Response Curves in Human Cone Vision. Journal of the Optical Society of America 50 (8): 780. ISSN 0030-3941. doi:10.1364/JOSA.50.000780.
- Bowmaker J K, Dartnall H J, Lythgoe J N and Mollon J D (1978). The visual pigments of rods and cones in the rhesus monkey, Macaca mulatta.. The Journal of Physiology 274 (1): 329–348. ISSN 00223751. doi:10.1113/jphysiol.1978.sp012151.
- Rushton W. A. H. (1963). A cone pigment in the protanope. The Journal of Physiology 168 (2): 345–359. ISSN 00223751. doi:10.1113/jphysiol.1963.sp007196.
- King-Smith P. E. (1973). The optical density of erythrolabe determined by a new method. The Journal of Physiology 230 (3): 551–560. ISSN 00223751. doi:10.1113/jphysiol.1973.sp010203.
- King-Smith P. E. (1973). The optical density of erythrolabe determined by retinal densitometry using the self-screening method. The Journal of Physiology 230 (3): 535–549. ISSN 00223751. doi:10.1113/jphysiol.1973.sp010202.
- Renner A.B., Knau H., Neitz M., Neitz J., and Werner J.S. (2005). Photopigment optical density of the human foveola and a paradoxical senescent increase outside the fovea. Visual Neuroscience 21 (06): 827–834. ISSN 0952-5238. doi:10.1017/S0952523804216030.
- Harwerth R.S. and Sperling H.G. (1975). Effects of intense visible radiation on the increment-threshold spectral sensitivity of the rhesus monkey eye. Vision Research 15 (11): 1193–1204. ISSN 00426989. doi:10.1016/0042-6989(75)90162-5.
- Vienot Francoise (2001). Retinal distributions of the macular pigment and the cone effective optical density from colour matches of real observers. Color Research & Application 26 (S1): S264–S268. ISSN 0361-2317. doi:10.1002/1520-6378(2001)26:1+<::AID-COL57>3.0.CO;2-3.
- Tornow R.P. and Stilling R. (1998). Variation in Sensitivity, Absorption and Density of the Central Rod Distribution with Eccentricity. Cells Tissues Organs 162 (2-3): 163–168. ISSN 1422-6405. doi:10.1159/000046482.
- Brindley, G.S. (1953). The effects on colour vision of adaptation to very bright lights. The Journal of Physiology 122 (2): 332–350. ISSN 00223751. doi:10.1113/jphysiol.1953.sp005003.
- Knowles A. and Dartnall H.J.A. (1977). The Photobiology of Vision. У Davson H. The Eye. Vol.2, part. 2. Academic Press. ISBN 978-0-12-206762-4.
- Fein Alan and Szuts Ete Zoltan (31 серпня 1982). Photoreceptors: Their Role in Vision. CUP Archive. ISBN 978-0-521-28684-8.
- Stiles W. S. and Crawford B. H. (1933). The Luminous Efficiency of Rays Entering the Eye Pupil at Different Points. Proceedings of the Royal Society B: Biological Sciences 112 (778): 428–450. ISSN 0962-8452. doi:10.1098/rspb.1933.0020.
- Goldmann, Hans (1942). Stiles-Crawford-Effekt. Ophthalmologica (German) 103 (4): 225–229. ISSN 1423-0267. doi:10.1159/000300022.
- Enoch Jay (1964). Physical properties of the retinal receptor and response of retinal receptors.. Psychological Bulletin 61 (4): 242–251. ISSN 0033-2909. doi:10.1037/h0047646.
- Snyder Allan W. and Pask Colin (1973). The Stiles-Crawford effect—explanation and consequences. Vision Research 13 (6): 1115–1137. ISSN 00426989. doi:10.1016/0042-6989(73)90148-X.
- Enoch, J.M., and Lakshminarayanan, V. (1991). Retinal Fibre Optics. У Charman W.N. Visual Optics and Instrumentation. Vol. 1 : Visual Optics and Instrumentation. CRC Press. с. 280–309. ISBN 978-0-8493-7501-9.
- Laties AM and Enoch JM (1971). An analysis of retinal receptor orientation. I. Angular relationship of neighboring photoreceptors.. Investigative ophthalmology 10 (1): 69–77. PMID 4992333.
- Bonds AB and MacLeod DI (1978). A displaced Stiles-Crawford effect associated with an eccentric pupil.. Investigative Ophthalmology & Visual Science 17 (8): 754–61. PMID 681135.
- Applegate RA and Bonds AB (1981). Induced movement of receptor alignment toward a new pupillary aperture.. Investigative Ophthalmology & Visual Science 21 (6): 869–72. PMID 7309438.
- Kono M., Enoch J.M., Strada E., Shih P., Srinivasan R., Lakshminarayanan V., Susilasate W. and Graham A. (2001). Stiles–Crawford effect of the first kind: assessment of photoreceptor alignments following dark patching. Vision Research 41 (1): 103–118. ISSN 00426989. doi:10.1016/S0042-6989(00)00228-5.
- Berendschot T., DeLint Peter Jaap and Norren Dirk van (2003). Fundus reflectance—historical and present ideas. Progress in Retinal and Eye Research 22 (2): 171–200. ISSN 13509462. doi:10.1016/S1350-9462(02)00060-5.
- Packer OS, Williams DR and Bensinger DG (1996). Photopigment transmittance imaging of the primate photoreceptor mosaic.. journal of neuroscience 16 (7): 2251–60. PMID 8601805.
- Sincich L.C. , Sabesan R., Tuten W.S. , Roorda A. and Harmening W.M. (19 грудня 2016). Chapter 3. Functional Imaging of Cone Photoreceptors. У Kremers J., Baraas R.C. and Marshall N.J. Human Color Vision. Springer Series in Vision Research, Volume 5. Springer. с. 71–104. ISBN 978-3-319-44978-4.
- Enoch J. M. (1961). Wave-Guide Modes in Retinal Receptors. Science 133 (3461): 1353–1354. ISSN 0036-8075. doi:10.1126/science.133.3461.1353.
- Enoch Jay M. (1960). Waveguide Modes: Are They Present, and What Is Their Possible Role in the Visual Mechanism?*. Journal of the Optical Society of America 50 (10): 1025. ISSN 0030-3941. doi:10.1364/JOSA.50.001025.
- Enoch Jay M. (1961). Nature of the Transmission of Energy in the Retinal Receptors. Journal of the Optical Society of America 51 (10): 1122. ISSN 0030-3941. doi:10.1364/JOSA.51.001122.
- Enoch Jay M. (1963). Optical Properties of the Retinal Receptors. Journal of the Optical Society of America 53 (1): 71. ISSN 0030-3941. doi:10.1364/JOSA.53.000071.
- Enoch Jay M. (1961). Visualization of Wave-Guide Modes in Retinal Receptors. American Journal of Ophthalmology 51 (5): 1107/235–1118/246. ISSN 00029394. doi:10.1016/0002-9394(61)91800-1.
- Marcos S. and Burns S. A. (1999). Cone spacing and waveguide properties from cone directionality measurements. Journal of the Optical Society of America A 16 (5): 995. ISSN 1084-7529. doi:10.1364/JOSAA.16.000995.
- Vohnsen B., Iglesias I. and Artal P. (2005). Guided light and diffraction model of human-eye photoreceptors. Journal of the Optical Society of America A 22 (11): 2318. ISSN 1084-7529. doi:10.1364/JOSAA.22.002318.
- Vohnsen Brian (2014). Directional sensitivity of the retina: A layered scattering model of outer-segment photoreceptor pigments. Biomedical Optics Express 5 (5): 1569. ISSN 2156-7085. doi:10.1364/BOE.5.001569.
- Brucke, Ersnt von (1843). Ueber den inneren Bau des Glaskörpers. Archiv für Anatomie, Physiologie und Wissenschaftliche Medicin (German) 11. Berlin: Müller, Joh. с. 345–348.
- B Brucke, Ersnt von (1844). Ueber die physiologische Bedeutung der stabförmigen Körper und der Zwillingszapfen in den Augen der Wirbelthiere. Archiv für Anatomie, Physiologie und wissenschaftliche Medicin (German). Berlin: Müller, Joh. с. 444–451.
- Brucke, Ersnt von (1847). Ueber das Leuchten der menschlichen Augen. Archiv für Anatomie, Physiologie und wissenschaftliche Medicin (German). Berlin: Muller, Joh. с. 225–227.
- Brucke, Ernst von (1847). Bemerkungen über die Bestimmung des specifischen Gewichtes der Milch. Archiv für Anatomie, Physiologie und wissenschaftliche Medicin (German). Berlin: Muller, Joh. с. 409–414.
- Brucke, Ernst von (1847). Nachtrag zu meinem Aufsatze über das Leuchten der Augen bei den Menschen. Archiv für Anatomie, Physiologie und wissenschaftliche Medicin (German) 14. Berlin: Muller, Joh. с. 479–480.
- Snyder, A.W. and Love, J. (30 листопада 1983). Optical Waveguide Theory. Springer. с. 734. ISBN 978-0-412-09950-2.
- Sidman R. L. (1957). The structure and concentration of solids in photoreceptor cells studied by refractometry and interference microscopy. Journal of Biophysical and Biochemical Cytology 3 (1): 15–30. ISSN 0021-9525. doi:10.1083/jcb.3.1.15.
- Burns Stephen A., Wu Shuang, He Ji Chang and Elsner Ann E. (1997). Variations in photoreceptor directionality across the central retina. Journal of the Optical Society of America A 14 (9): 2033. ISSN 1084-7529. doi:10.1364/JOSAA.14.002033.
- Gorrand Jean-Marie and Delori Francois (1995). A reflectometric technique for assessing photoreceptor alignment. Vision Research 35 (7): 999–1010. ISSN 00426989. doi:10.1016/0042-6989(94)00203-X.
- Francia G Toraldo di (1949). The Radiation Pattern of Retinal Receptors. Proceedings of the Physical Society. Section B 62 (7): 461–462. ISSN 0370-1301. doi:10.1088/0370-1301/62/7/108.
- Hannover, A. (1843). Mikroskopiske undersögelser af nervesystemet. Det Kongelige Danske videnskabernes selskabs naturvidenskabelige og mathematiske afhandlinger 10. Det Selskab. с. 1–112.
- Gorrand J.M. (2006). The directionality of photoreceptors in the human retina.. Bulletin de la Societe belge d'ophtalmologie (302): 215–29. PMID 17265800.
- van Hateren, J. Hans (1989). Photoreceptor Optics, Theory and Practice (Chapter 4). У Stavenga D.G., Hardie R.C. Facets of Vision. Springer Science & Business Media. с. 74–89. ISBN 978-3-642-74082-4.
- Snyder Allan W. and Pask Colin (1973). Waveguide modes and light absorption in photoreceptors. Vision Research 13 (12): 2605–2608. ISSN 00426989. doi:10.1016/0042-6989(73)90261-7.
- MENZEL RANDOLF and SNYDER ALLEN W. (1983). Photoreceptor Optics - Structure and Function Of Photoreceptors. У Hoppe W. , Lohmann W., Markl H. and Ziegler H. Biophysics. Springer. ISBN 978-3-642-68877-5.
- Levi, Dennis M. (3 березня 2011). Visual Acuity (Chapter 33). У Levin L.A. et al. Adler's Physiology of the Eye. Elsevier Health Sciences. с. 627–647. ISBN 0-323-08116-9.
- Williams David R. and Hofer Heidi (2004). 50. Formation and Acquisition of the Retinal Image. У Chalupa L.M. , Werner J.S. The Visual Neurosciences. MIT Press. с. 795–810. ISBN 978-0-262-03308-4.
- Yellott, John I. (15 січня 1990). The Photoreceptor Mosaic as an Image Sampling Device. У Committee on Vision, National Research Council. Advances in Photoreception: Proceedings of a Symposium on Frontiers of Visual Science. National Academies Press. с. 117–134. ISBN 978-0-309-54339-2.
- Bergmann, Carl (1858). Anatomisches und physiologisches über die Netzhaut des Auges. Zeitschrift für rationelle Medicin 2. Heidelberg- Leipzig. с. 83–108.
- D'Zmura, Michael (1996). Bergmann on Visual Resolution. Perception 25: 1223–1234.
- Helmholtz, Hermann von (1867). Handbuch der physiologischen Optik (German). Leipzig : Leopold Voss. с. 926.
- MacKay, David J. C. (25 вересня 2003). Information Theory, Inference and Learning Algorithms. Cambridge University Press. ISBN 978-0-521-64298-9.
- Goodman, Joseph W. (January 2005). Introduction to Fourier Optics. Roberts and Company Publishers. с. 491. ISBN 978-0-9747077-2-3.
- Marks II, Robert J (8 січня 2009). Handbook of Fourier Analysis & Its Applications. Oxford University Press. с. 687. ISBN 978-0-19-804430-7.
- Anderson S.J., Mullen K.T. and Hess R.F. (1991). Human peripheral spatial resolution for achromatic and chromatic stimuli: limits imposed by optical and retinal factors.. The Journal of Physiology 442 (1): 47–64. ISSN 00223751. doi:10.1113/jphysiol.1991.sp018781.
- Williams David R. and Coletta Nancy J. (1987). Cone spacing and the visual resolution limit. Journal of the Optical Society of America A 4 (8): 1514. ISSN 1084-7529. doi:10.1364/JOSAA.4.001514.
- Yellott John I. , Wandell Brian A. and Tom N. Cornsweet (1984). The Beginnings of Visual Perception: The Retinal Image and its Initial Encoding.. У Brookhart John Mills and Mountcastle Vernon B. Handbook of Physiology: A Critical, Comprehensive Presentation of Physiological Knowledge and Concepts. American Physiological Society. с. 257–316. ISBN 978-0-683-01108-1.
- Petersen Daniel P. and Middleton David (1962). Sampling and reconstruction of wave-number-limited functions in N-dimensional Euclidean spaces. Information and Control 5 (4): 279–323. ISSN 00199958. doi:10.1016/S0019-9958(62)90633-2.
- Packer O and Williams DR (11 липня 2003). Chapter 2. Light, the Retinal Image, and Photoreceptors. У Shevell S.K. The Science of Color (вид. 2). Elsevier. с. 41–102. ISBN 978-0-08-052322-4.
- Field David J. (1987). Relations between the statistics of natural images and the response properties of cortical cells. Journal of the Optical Society of America A 4 (12): 2379. ISSN 1084-7529. doi:10.1364/JOSAA.4.002379.
- Boreman, Glenn D. (2001). Modulation Transfer Function in Optical and Electro-optical Systems. SPIE Press. с. 110. ISBN 978-0-8194-4143-0.
- Yellott J. I. (1982). Spectral analysis of spatial sampling by photoreceptors: Topological disorder prevents aliasing. Vision Research 22 (9): 1205–1210. ISSN 00426989. doi:10.1016/0042-6989(82)90086-4.
- Osterberg, Gustav (1935). Topography of the Layer of Rods and Cones in the Human Retina. Acta ophthalmologica: Supple-mentum. Levin & Munksgaard. с. 102.
- Campbell F. W. and Gubisch R. W. (1966). Optical quality of the human eye. The Journal of Physiology 186 (3): 558–578. ISSN 00223751. doi:10.1113/jphysiol.1966.sp008056.
- Gubisch R. W. (1967). Optical Performance of the Human Eye. Journal of the Optical Society of America 57 (3): 407. ISSN 0030-3941. doi:10.1364/JOSA.57.000407.
- Jennings J.A.M. and Charman W.N. (1981). Off-axis image quality in the human eye. Vision Research 21 (4): 445–455. ISSN 00426989. doi:10.1016/0042-6989(81)90091-2.
- Galvin Susan J. and Williams David R. (1992). No aliasing at edges in normal viewing. Vision Research 32 (12): 2251–2259. ISSN 00426989. doi:10.1016/0042-6989(92)90089-2.
- Huck, F. O., Fales, C. L., Park, S. K., Samms, R. W. and Self, M. O. (December 1983). Information theory analysis of sensor-array imaging systems for computer vision. У Oosterlinck André. Applications of digital image processing: April 19-22, 1983, Geneva, Switzerland. International Society for Optical Engineering. с. 82–99. ISBN 978-0-89252-432-7.
- Thibos L. N. (1987). Calculation of the influence of lateral chromatic aberration on image quality across the visual field. Journal of the Optical Society of America A 4 (8): 1673. ISSN 1084-7529. doi:10.1364/JOSAA.4.001673.
- Yellott John I. (1984). Image sampling properties of photoreceptors: A reply to Miller and Bernard. Vision Research 24 (3): 281–282. ISSN 00426989. doi:10.1016/0042-6989(84)90132-9.
- Snyder A., Bossomaier T. and Hughes A. (1986). Optical image quality and the cone mosaic. Science 231 (4737): 499–501. ISSN 0036-8075. doi:10.1126/science.3941914.
- Hsu Andrew, Smith Robert G., Buchsbaum Gershon and Sterling Peter (2000). Cost of cone coupling to trichromacy in primate fovea. Journal of the Optical Society of America A 17 (3): 635. ISSN 1084-7529. doi:10.1364/JOSAA.17.000635.
- Ruderman Daniel L. and Bialek William (1992). Seeing Beyond the Nyquist Limit. Neural Computation 4 (5): 682–690. ISSN 0899-7667. doi:10.1162/neco.1992.4.5.682.
- Williams D.R. (1985). Aliasing in human foveal vision. Vision Research 25 (2): 195–205. ISSN 00426989. doi:10.1016/0042-6989(85)90113-0.