Хронологія еволюції людини
Хронологія людської еволюції включає до себе основні події розвитку людського виду, людини розумної, і еволюції наших предків. До неї входять короткі описи деяких видів, родів і вищих таксономічних ранґів, які розглядаються сьогодні як можливі предки сучасних людей.
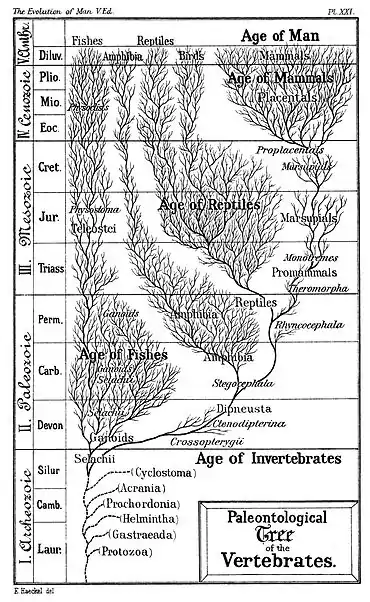
Цей графік ґрунтується на дослідженнях з антропології, палеонтології, біології розвитку, морфології, а також від анатомічних і генетичних даних. Він не розглядає походження життя, дослідженням чого займається абіогенез, а являє собою одну з можливих ліній еволюційного походження видів, що в кінцевому підсумку призвело до появи сучасних людей.
Таксономія людини розумної
Одна з декількох можливих ліній походження людини розумної показана нижче.
| Ранґ | Наукова назва | Поширена назва | Мільйони років тому |
|---|---|---|---|
| Домен | Eukaryota | Ядерні клітини | 2,100 |
| Царство | Animalia | Тварини | 590 |
| Тип | Chordata | Хордові | 530 |
| Підтип | Vertebrata | Хребетні | 505 |
| Надклас | Tetrapoda | Чотириногі | 395 |
| без ранґу | Amniota | Амніоти — усі чотириногі, у яких утворюються зародкові листки | 340 |
| Клада | Synapsida | Звіроящери | 308 |
| Клас | Mammalia | Ссавці | 220 |
| Підклас | Theria | Звірі | 160 |
| Інфраклас | Eutheria | Вищі звірі (плацентарні) | 125 |
| Магноряд | Boreoeutheria | Включає лавразіотерієв і еуархонтоглірів | |
| Надряд | Euarchontoglira | «Дійсні пращури»: примати, кагуанові, тупаєподібні, гризуни і зайцеподібні | 100 |
| Грандряд | Euarchonta | Примати, кагуанові і тупаєподібні | |
| Мірряд | Primatomorpha | Примати і кагуанові | 79.6 |
| Ряд | Primates | Примати | 75 |
| Підряд | Haplorrhini | Сухоносі примати: гоміноїди, мавпи і довгоп'яти | 63 |
| Інфраряд | Simiiformes | Вищі примати, або мавповиді | 40 |
| Парворяд | Catarrhini | Вузьконосі примати — мавпи Старого Світу | 30 |
| Надродина | Hominoidea | Людинові і гібонові | 28 |
| Родина | Hominidae | Людинові: люди, шимпанзе, горили і орангутани | 15 |
| Підродина | Homininae | Люди, шимпанзе і горили (африканські мавпи) | 8 |
| Триба | Hominini | Роди Homo, Pan і ще кілька викопних родів | 5.8 |
| Підтриба | Hominina | Рід Homo включаючи людських родичів і предків після розділення від роду Pan | 2.5 |
| Рід | Homo | Люди | 2.5 |
| Підрід | Homo | Археоантропи | 0.6 |
| Вид | Homo sapiens | Анатомічно сучасна людина | 0.2 |
| Підвид | Homo sapiens sapiens | Існуючі сучасні люди | 0.07? |
Хронологія
Перші живі істоти
| Дата | Події |
|---|---|
| 4100 млн (мільйонів років тому) |
З'являється раннє життя. |
| 3900 млн | З'являються перші прокаріоти серед клітин. |
| 3500 млн | Поява перших фотосинтезуючих організмів і, отже, поява великої кількості атмосферного кисню на Землі. |
| 2500 млн | З'являються організми, що використовують кисень. Близько 2,4 млрд років тому, під час Кисневої катастрофи, анаеробні форми життя були майже повністю витиснути кисневими. |
| 2100 млн | З'являються більш складні клітини: еукаріоти. |
| 1200 млн | Розвивається статеве розмноження, що призводить до швидшого еволюції[1], де гени змішуються в кожному поколінні, що дає велику варіацію для подальшого відбору. |
| 900 млн |
Комірцеві джгутиконосці можуть бути предками всього тваринного світу, і, зокрема, вони можуть бути прямими предками губок.[2] Протогубки (рід хоанофлагелятів) є кращими живими прикладами того, як міг виглядати преддок всіх тварин. Вони живуть в колоніях і демонструють примітивний рівень клітинної спеціалізації для різних завдань. |
| 600 млн | Вважається, що найранішою багатоклітинною твариною була губка.
Губки відносяться до найпростіших тварин з частково диференційованими тканинами. Губки (Porifera) є філогенетично найстарішим типом тварин, існуючий сьогодні. |
| 580 млн | Можливо, рух у багатоклітинних тварин почався з кнідарій. Майже всі кнідарії володіють нервами і м'язами. Оскільки вони є найпростішими тваринами, що володіють ними, їх безпосередні предки були, ймовірно, першими тваринами, які використовують разом нерви і м'язи. Кнідаріі також є першими тваринами з реальним тілом певної форми і форми. Воно мають радіальну симетрію. У цей час з'явилися перші очі. |
| 550 млн | Плоскі черви — найдавніші з відомих тварин, у яких є мозок, і найпростіші існуючі тварини, що мають двосторонню симетрію. Вони також є простішими тваринами з органами, які формуються з трьох зародкових шарів. Найвідоміші знайдені скам'янілості тварин були морськими видами часів кембрійського вибуху. |
| 540 млн | Кишководишні вважаються більш вузькоспеціалізованими і розвиненими червоподібними, ніж інші аналогічні форми. Вони мають систему кровообігу з серцем, яке також функціонує як нирка. Кишководишні мають зяброву структуру, використовувану для дихання, структуру, подібну структурі примітивних риб. Іноді кажуть, що кишководишні є сполучною ланкою між хребетними і безхребетними.[джерело?] |


Хордові
| Date | Event |
|---|---|
| 530 млн | Пікайя — знаковий предок сучасних хордових і хребетних.[3] Іншими найранішими хордовими були Myllokunmingia fengjiaoa,[4] Haikouella lanceolata,[5] і Haikouichthys ercaicunensis.[6]
Ланцетник, який все ще живе сьогодні, зберігає деякі характеристики примітивних хордових, чим нагадує пікаю. Конодонти — відомий ранній (495 млн років тому і пізніше) тип викопного хордового; вони мали своєрідні зуби як у вугрів, широкі очі, плавці з променями, шевронні м'язи і хорду. |
| 505 млн |
З'являються перші хребетні: остракодерми, безщелепні риби, з якими пов'язані сучасні міноги і міксини. Haikouichthys і Myllokunmingia є прикладами цих безщелепних риб. Вони не мали щелепи і їх внутрішні скелети були хрящовими. У них не було парних (грудних і тазових) плавців, як у просунутішої риби. Вони були попередниками кісткових риб.[7] |
| 480 млн |
Панцирні риби (Placodermi) були одними з перших щелепних (Gnathostomata), їх щелепи розвивалися з першої зябрової дуги.[8] Голова і груди плакодерми були покриті зчленованими броньованими пластинами, а інша частина тіла була лускатою або оголеною. Проте, знайдені скам'янілості вказують, що вони не залишили нащадків після закінчення девону і менш тісно пов'язані з живими кістковими рибами, ніж акули.[джерело?] |
| 410 млн | Вперше з'являються целакантоподібні;[9] Передбачалося, що цей ряд тварин давно вимер, поки живі екземпляри єдиного вцілілого виду не були виявлені 1938 року. Його часто називають живим викопним. |



Чотириногі
| Дата | Події |
|---|---|
| 390 млн |
Деякі прісноводні лопатепері риби (Sarcopterygii) розвивають ноги і дають початок тетраподам. Перші чотириногі розвивалися в дрібних і болотистих прісноводних місцепроживаннях. Примітивні чотириногі розвинулися з лопатоперих риб, з дводольним мозком у сплюснутому черепі, широким ротом і короткою мордою, чиї звернені догори очі вказують, що вони були мешканцями дна, і які вже розробили пристосовані м'ясисті плавці з кістками («Живий викопний» латимерія — родинна з лопатоперими риба, але без цих мілководних пристосувань). Тетраподні риби використовували свої плавці як лопаті у мілководних місцях проживання, забитих рослинами і детритом. Універсальні тетраподні характеристики передніх кінцівок, які згинаються назад в ліктях, і задніх кінцівок, що згинаються вперед в коліні, можна з певністю простежити в ранніх чотириногих, що жили на мілководді.[10] Пандеріхтіс — 90–130 сантиметрова довга риба пізнього девонського періоду (380 млн років тому). В неї була велика тетраподоподібна голова. Panderichthys показує особливості, перехідні між лотоперими рибами і ранніми чотириногими. Відбитки слідів, зроблені чимось, що нагадує кінцівки іхтіостеги, були зроблені 390 млн років тому в морських припливних відкладеннях Польщі. Це свідчить про те, що еволюція тетраподів є давнішою, ніж датовані скам'янілості Пандеріхтіса до Іхтіостеги. Дводишні зберегли деякі характеристики ранніх тетраподів. Одним із прикладів є рогозуб. |
| 375 млн |
Тіктаалік — рід лопатоперих риб з пізнього девону з багатьма тетраподобними ознаками, що вказує на його чіткий зв'язок між Пандеріхтісом і Іхтіостегою. |
| 365 млн |
Акантостега — вимерла амфібія, серед перших тварин, у якій були пізнавані кінцівки. Це кандидатка на перших хребетних, що змогли вийти на суходіл. Їй не вистачало зап'ясть і вона зазвичай погано адаптувалася до життя на суші. Кінцівки не могли підтримувати вагу тварини. Акантостега мала як легені, так і зябра, що також вказує на те, що вона була сполучною ланкою між лопатоперими рибами і наземними хребетними. Іхтіостега була найранішим чотириногим. Будучи однією з перших тварин з ногами, руками і кістками пальців, іхтіостега розглядається як гібрид між рибою і земноводним. Іхтіостега мала кінцівки, але вони, ймовірно, не використовувалися для ходьби. Вона, можливо, проводила дуже мало часу поза водою і використовувала свої лапи, щоб просуватися через бруд.[11] Земноводні були першими чотириногими тваринами, що розвинули повноцінні легені, котрі, можливо, еволюціонували від гінерпетона 360 млн років тому. Земноводні, що живуть сьогодні, досі зберігають багато характеристик ранніх чотириногих. |
| 300 млн |  Гілоном (Hylonomus) Від земноводних з'явилися перші плазуни: Гілоном — найраніший з відомих плазунів. Він був 20 см завдовжки (включаючи хвіст) і, ймовірно, виглядав би досить схожим на сучасних ящірок.Він мав невеличкі гострі зуби та, вірогідно, їв багатоніжок і ранніх комах. Це попередник пізніших амніотів і звіроподібних плазунів. Вперше тут розвивається альфа кератин, який знаходиться в пазурах сучасних ящірок і птахів, а також у волоссі ссавців.[12] Еволюція амніотичного яйця призводить до появи амніотів, плазунів, які можуть розмножуватися на землі і відкладати яйця на суші, тому їм більше не потрібно було повертатися до води для розмноження. Ця адаптація дала їм можливість вперше заселитися вглиб материка. В плазунів дужче розвинулася нервова система, у порівнянні з земноводними. В них дванадцять пар черепних нервів. |




Ссавці
| Дата | Події |
|---|---|
| 256 млн | Незабаром після появи перших плазунів, дві гілки розкололися. Одна гілка — зауропсиди, з яких походять сучасні плазуни і птиці. Інша гілка — синапсиди, з якої походять сучасні ссавці. В обох були скроневі фенестри, пара отворів в черепах за очима, які використовувалися, щоб збільшити простір для м'язів щелепи. У синапсидів був один отвір з кожного боку, а у діапсидів два.
Найранішими звіроподібними плазунами були пелікозаври. Вони були першими тваринами, в яких були черепні фенестри. Пелікозаври ще не були терапсидами, але невдовзі вони породили їх. Терапсиди були прямими пращурами ссавців. Терапсиди мали все звіроподібніші черепні фенестри, ніж пелікозаври, їх зуби показують послідовнішу диференціацію, а пізніші форми розвили вторинне піднебіння. Вторинне піднебіння дозволяє тварині їсти і дихати одночасно і є ознакою активнішого, можливо, теплокровного способу життя.[13] |
| 220 млн |
В однієї підгрупи терапсидів, цинодонтів, усе більше розвивалися звіроподібні ознаки. Щелепи цинодонтів нагадують сучасні щелепи ссавців. Досить імовірно, що ця група тварин містила вид, який став прямим пращуром всіх сучасних ссавців.[14] </ref> |
| 220 млн |  Репеномам (Repenomamus) Першими ссавцями були еуцинодонти (інфраряд цинодонтів). Більшість ранніх ссавців були дрібними землерийними тваринами, які харчувалися комахами. Хоча немає ніяких доказів в знайдених скам'янілостях, цілком вірогідно, що у цих тварин була постійна температура тіла і молочні залози для вигодовування потомства. Область неокортексу мозку спочатку еволюціонувала саме від ссавців і тому унікальна тільки для них. Однопрохідні є яйцекладущей групою ссавців, представленої серед сучасних тварин качкодзьобом і єхиднами. Недавнє секвенування генома у качкодзьоба вказує на те, що його статеві гени ближче до пташиних, ніж до живородних ссавців (звірів). Порівнюючи це з іншими ссавцями, можна зробити висновок у тому, що перші ссавці, які отримали статеву диференціацію внаслідок існування або відсутності гена SRY (виявленого в Y-хромосомі), еволюціонували після розщеплення монотріальной лінії. |
| 160 млн |  Juramaia sinensis Juramaia sinensis[15] — найраніший з відомих евферийских ссавців. |
| 100 млн | Останній загальний предок мишей і людей (початок клади еуархонтоглірів). |

Примати
| Дата | Події |
|---|---|
| 85–65 млн |  Плезіадап (Plesiadapis)  Carpolestes simpsoni Група маленьких, нічних і деревних комахоїдних ссавців, яких називають еуархонтами, починається з видоутворення, яке призведе до приматів, тупаєподібних і шерстокрилих. Приматоморфи — надряд еуархонтів, до якої входять примати і вимерлі плезіадапоподібні. Одним з найраніших протоприматів був Плезіадап. Він все ще зберігав кігті і мав очі, розташовані з боків голови. Через це, вони були швидші на землі, ніж на верхів'ї дерев, але також стали витрачати багато часу на нижніх гілках дерев, де поїдали плоди і листя. Найімовірніше, до плезіадапоподібних входив вид, який став пращуром всім приматам.[16] Одним з останніх плезіадапоподібних був Carpolestes simpsoni. В нього були кінцівки, пристосовані до хапання, але очі все ще були з боків. |
| 63 млн | Примати розійшлися в підряди мокроносих і сухоносих. До мокроносих приматів увійшли більшість напівмавп; сучасними прикладами є лемури і лорі. Сухоносі примати включають до себе сучасних довгоп'ятів і мавп. Одним з найраніших сухоносих приматів був рід Теярдіна, денна істота мишачого розміу з маленькими очима. Метаболізм сухоносих втратив здатність виробляти свій власний вітамін C. Це означало, що вони і всі їх нащадки повинні будуть включати фрукти до свого раціону, де можна отримати необхідний вітамін С. |
| 30 млн |
Сухоносі розділилися на широконосих і вузьконосих приматів. Широконосі, або мавпи Нового Світу, мають чіпкі хвости, а самці — дальтоніки. Вони, можливо, мігрували до Південної Америки на плоту з рослинності через відносно вузький Атлантичний океан (близько 700 км). Вузьконосі примати залишилися переважно в Африці, коли два континенти розійшлися. Можливими ранніми пращурами вузьконосих були Єгиптопітеки і Саадани. |
| 25 млн |
Вузьконосі розпалися на дві надродини: мавпуваті (Cercopithecoidea) і людинуваті (Hominoidea). У цей період має своє генетичне походження наш трихроматичний (кольоровий) зір. Проконсул був раннім родом вузьконосих приматів. У ньогобула суміш ознак мавпуватих і людинуватих характеристик. У мавпячі особливості проконсула належали тонка зубна емаль, легка статура з вузькою грудною кліткою і короткими передніми кінцівками, та деревний чотириногий спосіб життя. Його людиноподібні особливості були відсутність хвоста, людиноподібні лікті і дещо більший мозок щодо розміру тіла. Африканський проконсул є можливим пращуром як вищих, так і малих мавп, включаючи людей. |


Гомініди
| Дата | Події |
|---|---|
| 15 млн | Гомініди (великі мавпи) відділяються від пращурів гібонів (малих мавп). |
| 13 млн | Пращури гомінідів відділяються від пращурів орангутанів.[17]
Каталонський піеролапітек вважається спільним предком людей і інших людиноподібних мавп або, принаймні, видом, що наближує нас до спільного предка ближче, ніж будь-яке попереднє викопне відкриття. В нього були спеціальні пристосування для лазіння по деревах, як і сучасних людей і інших вищих мавп: широка пласка грудна клітка, жорстка нижня частина хребта, гнучкі зап'ястя і лопатки, що лежать уздовж його спини. |
| 10 млн | Родовід, який нині представлений людьми і родом Пан (шимпанзе звичайний і бонобо), виділяється від пращурів горил. |
| 7 млн |
Гомініна, підтриба гомінінів, відокремлюється від пращурів шимпанзе. І у шимпанзе, і у людей є гортань, яка переміщається протягом перших двох років життя на місце між горлом і легенів, вказуючи, що їх спільні предки мали цю ж функцію, як одну з передумов для вокалізації мовлення у людей. Останній загальний пращур людей і шимпанзе жив за часів Сахельантропа бл. 7 млн років тому, та іноді стверджують, що саме S. tchadensis був їх останнім спільним предком, але точних підтверджень наразі не виявлено. Найраніший відомий представник з пращурів людської лінії після поділу з лінією шимпанзе був Orrorin tugenensis («Людина тисячоліття», Кенія; бл. 6 млн років тому). |
| 4.4 млн |
Ардипітек є або може бути дуже раннім родом гомінінів. У літературі описані два види: A. ramidus, який жив близько 4,4 мільйонів років тому[18] в ранньому Пліоцені, і A. kadabba, датований приблизно 5,6 мільйонам років[19] (пізній Міоцен). A. ramidus мав малий мозок потужністю між 300 і 350 см3. Цей мозок приблизно такого ж розміру, як мозок сучасних бонобо і самиць звичайного шимпанзе; він набагато менше мозку австралопітеків, таких як Люсі (400—550 см3), і ледве більш п'ятої величини сучасного мозку Людини розумної. Ардипітек був деревним, тобто жив переважно в лісі, де конкурував з іншими лісовими тваринами за їжу, без сумніву, включаючи пращура нинішнього шимпанзе. Ардипітек був, напевно, двоногим, про що свідчить його чашоподібна форма тазу, кут його великого потиличного отвору і його тонкі кісточки зап'ястя, хоча його ноги досі були пристосовані для хапання, а не для ходьби на великі відстані. |
| 3.6 млн |
Представник афарського австралопітека залишив схожі на людські сліди на вулканічному попелі в Лаетолі, Кенія (Північна Танзанія), що є переконливим свідченням безперервного біпедалізму. Австралопітек афарський жив між 3,9 і 2,9 млн років тому і вважається одним з ранніх гомінінів — тих видів, які розвивалися і становили лінію найближчих родичів Людини після поділу з лінією шимпанзе. Вважається, що А. афарський був пращуром як роду австралопітеків, так і роду людей. У порівнянні з сучасними і зниклими людиноподібними мавпами, у А. афарського скоротилися ікла і моляри, хоча вони були все ще відносно більше, ніж у сучасних людей. А. афарський також мав відносно невеликий розмір мозку (380—430 cm³) і прогнатичне (тобто, що виступає вперед) обличчя. Австралопітеки були знайдені в саванах; ймовірно, вони почали включати до свого раціону м'ясо. Аналіз нижніх хребців африканського австралопітека показує, що ці кістки змінилися у жінок так, щоб вони могли підтримувати двоногість навіть під час вагітності. |
| 3.5–3.3 млн | Кеніантроп платіопс, можливий пращур Homo, з'являється з роду австралопітеків. Свідомо майстрував кам'яні знаряддя.[20] |
| 3 млн | Двоногі австралопітеки (рід підтриби Hominina) еволюціонують у саванах Африки, на яких полює Динофеліс. Втрата волосся на тілі відбувається від 3 до 2 млн років тому, паралельно з розвитком повної двуногості. |

Людина
| Дата | Події |
|---|---|
| 2.8 млн |
Рід людей (Homo) з'являється в Східній Африці; разом з більшістю австралопітеків вони вважаються першими гомінінінами, тобто, вони позначаються (деякими) так, як ті самі ранні люди і людські родичі або пращури, які відійшли від лінії Пан, шимпанзе. Інші вважають рід Пан гомінінами, і, можливо, взагалі першими гомінінами. Поява складних кам'яних знарядь відзначає початок Нижнього Палеоліту. З'являється Людина уміла — перший, або один з перших, гомінін, який освоює технологію кам'яних інструментів. Інструменти кам'яного знаряддя також знайдені разом з Австралопітеком гарі, котрий датований трохи ранішим періодом.. Людина уміла, хоча і значно відрізняється анатомією і фізіологією, вважається родоначальником Людини працюючої, або африканської Людини прямоходячої; але також відомо, що він співіснував з Людиною прямоходячою протягом приблизно півмільйона років. |
| 1.8 млн |
В Африці розвивається Людина прямоходяча. Вона матиме разючу подібність з сучасними людьми, але мозок буде складати біля 74 відсотків від розміру сучасної людини. Її лоб менш похилий, ніж у Людини умілої, і зуби менші. Людина працююча, відома як африканська Людина прямоходяча, і інші види гомінін, такі як Грузинська людина, Пекінська людина, Гейдельберзька людина, часто поміщаються під загальну назву виду Л. прямоходячої.[21] Починаючи з Грузинської людини, знайденого в республіці Грузії віком 1,8 млн років, таз і хребет стали схожіши на людські, що дозволило б Л. грузинської долати дуже великі відстані і слідувати за стадами копитних тварин; це найстаріша скам'янілість гомініна, знайдена за межами Африки. Освоєння вогню первісними людьми відбулося 1,5 млн років тому Людиною працюючою. Вона вже досягала висоти до 1,9 метрів. Формування темної шкіри, яка пов'язана з втратою волосся на тілі у людських пращурів, завершується 1,2 млн років тому. Пекінська людина вперше з'являється в Азії близько 700 тис. років тому, але, відповідно до теорії африканського походження людей, вона не могла бути пращуром сучасних людей, а скоріше за все була відгалуженим двоюрідним видом Л. прямоходячої. Гейдельберзька людина був дуже великим гомініном, який розробив досконаліши ріжучі знаряддя і, можливо, полював на велику дичину, таку як коні. |
| 1.2 млн | Людина-попередник може бути спільним пращуром людей і неандертальців.[22][23] Наразі оцінюється, що у людей є приблизно 20 000-25 000 генів і частка в 99 % їх ДНК, що збігається з ДНК вимерлих неандертальців[24], а 95-99 % ДНК — з найближчим живим еволюційним родичем людини, шимпанзе.[25][26] Було встановлено, що людський варіант гену FOXP2 (пов'язаний з контролем мови) ідентичний і у неандертальців.[27] |
| 600 тис. (тисячу років тому) |
Три півтораметрові Гейдельберзькі людини залишили сліди в вулканічному попелі, затверділому в Італії. Л. гейдельберзька може бути спільним предком людей і неандертальців. Вона морфологічно дуже схожа на Л. прямоходячу, але Л. гейдельберзька мала більший череп, приблизно 93 % від розміру Л. розумної. Голотип виду був високим, 1,8 м заввишки і м'язистіша, ніж сучасні люди. Початок середнього палеоліту. |
| 500 тис. | Розходження ліній неандертальців і Денисівської людини від загального предка.[28] |
| 200 тис. | Місця Oмо1 и Oмо2 (річка Омо, Ефіопія) дають найраніші докази скам'янілостей анатомічно сучасної людини.[29]
За дослідження 2015 року, гіпотетичний Y-хромосомний Адам, за оцінками, жив у Східній Африці близько 250 тисяч років тому. Він був би останнім спільним предком, з якого походять всі чоловічі людські Y-хромосоми.[30] |
| 160 тис. | Людина розумна (Людина розумна найстаріша) в Ефіопії, річці Аваш (недалеко від нинішнього села Херто) практикує обрядові ритуали. Потенційні ранні свідоцтва анатомії і аспекти поведінкової сучасності узгоджуються з гіпотезою безперервності, включаючи використання червоної вохри і лову риби.[31]
Передбачувана мітохондріальна Єва, за оцінками, проживала в Східній Африці від 99 до 200 тисяч років тому. |
| 90 тис. | Поява мітохондріальної гаплогрупи (мт-гаплогрупа) L2. |
| 60 тис. | Поява mt-гаплогрупп M і N, які брали участь в міграції з Африки. Л. розумна, що залишила Африку в цій хвилі, можливо, схрещувалася з неандертальцями, з якими вона стикалася.[32][33] |
| 50 тис. | Поведінкова сучасність розвивається, відповідно до теорії «великого стрибка уперед».[34]
Люди мігрують з Африки як єдине населення.[35] У наступному тисячолітті нащадки цього населення мігрують в Індію та Азію. Мутація M168 (переноситься усіма неафриканськими чоловіками). Початок верхнього палеоліту. Поява мт-гаплогруп U і K. |
| 40 тис. | Міграція до Австралії[36] і Європи, в якій розвивається Кроманьйонець. |
| 25–40 тис. | Незалежна лінія неандертальців вимирає. Поява Y-гаплогрупи R2 та мт-гаплогрупи J і X. |
| 10–20 тис. | Початок мезоліту / голоцену. Поява Y-гаплогрупи R1а та мт-гаплогрупи V і T. Еволюція світлої шкіри у європейців (SLC24A5).[38][39] Флореська людина вимирає, залишаючи Людину розумну як єдиний живий вид роду Людини. |
Див. також
Примітки
- «Експерименти з сексом були дуже важкими, — сказав Ґодард. — Під час експерименту потрібно тримати все інше постійним, крім аспекту інтересу. Це означає, що ніякі вищі організми не можуть бути використані, оскільки їм доводиться схрещуватися для розмноження і, отже, не забезпечувати безстатевого контролю.
Замість цього Ґодард і його колеги звернулися до одноклітинному організму — дріжджів, щоб перевірити, що секс дозволяє популяціям адаптуватися до нових умов швидше, ніж безстатеві популяції.» Sex Speeds Up Evolution, Study Finds (URL accessed on January 9, 2005) - «Proterospongia — рідкісний прісноводний найпростіші, колоніальний член Choanoflagellata.» «Сама Proterospongia не є пращуром губок. Проте, вона служить корисною моделлю для того, що може бути пращуром губок і інших метазоїв.» http://www.ucmp.berkeley.edu/protista/proterospongia.html Berkeley University
- «Вочевидь, у хребетних були пращури, що жили в кембрії, але вони ймовірно були безхребетними попередниками справжніх хребетних — протохордатів. Пікайя була у великій мірі просунутої для рівня найстарішого викопного протохордата.» Річард Докінз 2004 The Ancestor's Tale сторінка 289, ISBN 0-618-00583-8
- Shu, D. G.; Luo, H. L.; Conway Morris, S.; Zhang, X. L.; Hu, S. X.; Chen, L.; Han, J.; Zhu, M.; Li, Y.; Chen, L. Z. (1999). Lower Cambrian vertebrates from south China. Nature 402 (6757): 42–46. Bibcode:1999Natur.402...42S. doi:10.1038/46965.
- Chen, J. Y.; Huang, D. Y.; Li, C. W. (1999). An early Cambrian craniate-like chordate. Nature 402 (6761): 518–522. Bibcode:1999Natur.402..518C. doi:10.1038/990080.
- Shu, D. G.; Morris, S. C.; Han, J.; Zhang, Z. F.; Yasui, K.; Janvier, P.; Chen, L.; Zhang, X. L.; Liu, J. N.; Li, Y.; Liu, H. -Q. (2003). Head and backbone of the Early Cambrian vertebrate Haikouichthys. Nature 421 (6922): 526–529. Bibcode:2003Natur.421..526S. PMID 12556891. doi:10.1038/nature01264.
- Цим першим хребетним не вистачало щелеп, як у нині живих міксин і міног. Щелепні хребетні з'явилися 100 мільйонів років тому, у силуру. http://www.ucmp.berkeley.edu/vertebrates/vertintro.html Berkeley University
- «Кістки першої зябрової дуги стали верхніми і нижніми щелепами.» (Зображення)
- Скам'яніла щелепа целеканта, знайдена в шарі віком 410 млн років, котра було зібрана близько Бучана в Вікторії, Австралійський східний Гіпсленд, наразі є найдавнішим знайденим екземпляром целекантів; йому було присвоєно ім'я «Eoactinistia foreyi», коли був опублікований у вересні 2006 року.
- «Дводишні, як вважають, є найближчими живими родичами чотириногих і мають з ними низку важливих характеристик. Серед цих ознак — зубна емаль, відділення легеневого кровотоку від кровотоку тілесного, розташування кісток черепа і наявність чотирьох кінцівок однакового розміру з тим же положенням і структурою, що і чотири ноги тетраподів.» http://www.ucmp.berkeley.edu/vertebrates/sarco/dipnoi.html Berkeley University
- «Чи розділяють земноводні загального пращура з рептиліями і нами?» «Ці можливі перехідні скам'янілості були добре вивчені, серед них акантостега, яка, здається, була повністю водною, і іхтіостега» Річард Докінз 2004 The Ancestor's Tale сторінка 250, ISBN 0-618-00583-8
- Eckhart L, Valle LD, Jaeger K; Valle; Jaeger; Ballaun; Szabo; Nardi; Buchberger; Hermann; Alibardi; Tschachler (Листопад 2008). Identification of reptilian genes encoding hair keratin-like proteins suggests a new scenario for the evolutionary origin of hair. Proceedings of the National Academy of Sciences of the United States of America 105 (47): 18419–23. Bibcode:2008PNAS..10518419E. PMC 2587626. PMID 19001262. doi:10.1073/pnas.0805154105.
- «У багатьох відносинах пелікозаври є проміжною ланкою між плазунами і ссавцями» http://www.ucmp.berkeley.edu/synapsids/pelycosaurs.html Berkeley University
- «Тринаксодона, як і будь-яку скам'янілість, слід розглядати як двоюрідного брата нашого пращура, а не самого пращура. Він належав до звіроподібних рептилій, названими цинодонтами. Цинодонти були настільки схожі на ссавців, що принадно назвати їх ссавцями. Але кого це хвилює, як ми їх називаємо? Вони майже ідеальна проміжна ланка.» Річард Докінз 2004 The Ancestor's Tale сторінка 211, ISBN 0-618-00583-8
- Luo, ZX; Yuan, CX; Meng, QJ; Ji, Q (Серпень 2011). A Jurassic eutherian mammal and divergence of marsupials and placentals. Nature 476: 442–445. PMID 21866158. doi:10.1038/nature10291.
- «Скам'янілості, які могли б допомогти нам відновити те, що було в Concestor 8, включає велику групу плезіадапоподібних. Вони жили приблизно в потрібний час, і у них є багато якостей, яких ви очікуєте від великого пращура всіх приматів» Річард Докінз 2004 The Ancestor's Tale page 136, ISBN 0-618-00583-
- Raauma, Ryan; Sternera, K (2005). Catarrhine primate divergence dates estimated from complete mitochondrial genomes (PDF). Journal of Human Evolution 48: 237–257. PMID 15737392. doi:10.1016/j.jhevol.2004.11.007.
- Perlman, David (12 липня 2001). Fossils From Ethiopia May Be Earliest Human Ancestor. National Geographic News. Процитовано Липень 2009. «Another co-author is Tim D. White, a paleoanthropologist at UC-Berkeley who in 1994 discovered a pre-human fossil, named Ardipithecus ramidus, that was then the oldest known, at 4.4 million years.»
- White, Tim D.; Asfaw, Berhane; Beyene, Yonas; Haile-Selassie, Yohannes; Lovejoy, C. Owen; Suwa, Gen; WoldeGabriel, Giday (2009). Ardipithecus ramidus and the Paleobiology of Early Hominids.. Science 326 (5949): 75–86. Bibcode:2009Sci...326...75W. PMID 19810190. doi:10.1126/science.1175802.
- Harmand S et al. ″3.3-million-year-old stone tools from Lomekwi 3, West Turkana, Kenya.″ Nature 521(7552) http://www.nature.com/nature/journal/v521/n7552/full/nature14464.html
- NOVA: Becoming Human Part 2 http://video.pbs.org/video/1319997127/
- J. M. Bermúdez de Castro, et al. A Hominid from the Lower Pleistocene of Atapuerca, Spain: Possible Ancestor to Neandertals and Modern Humans. Архівовано 3 грудня 2010 у Wayback Machine. Science 1997 May 30; 276: 1392—1395.
- Green, R. E.; Krause, J; Ptak, S. E.; Briggs, A. W.; Ronan, M. T.; Simons, J. F. (2006). Analysis of one million base pairs of Neanderthal DNA. Nature 444: 330–336. PMID 17108958. doi:10.1038/nature05336.
- «Рубін також сказав, що аналіз досі припускає, що людська і неандертальська ДНК приблизно на 99,5-99,9 % індентичні.» Neanderthal bone gives DNA clues (URL accessed on November 16, 2006)
- «Висновок: старе спостереження, де ми поділяємо 98,5 % нашої послідовності ДНК з шимпанзе, ймовірно, помилкове. » Britten, R.J. (2002). Divergence between samples of chimpanzee and human DNA sequences is 5%, counting indels. PNAS 99 (21): 13633–5. Bibcode:2002PNAS...9913633B. PMC 129726. PMID 12368483. doi:10.1073/pnas.172510699.
- «…з трьох мільярдів символів, що складають людський геном, тільки 15 мільйонів — менше 1 відсотка — змінилися за приблизно шість мільйонів років після того як лінії людини і шимпанзе розійшлися.» Pollard, K.S. (2009). What makes us human?. Scientific American. 300–5 (5): 44–49. doi:10.1038/scientificamerican0509-44.
- Krause J; Lalueza-Fox C; Orlando L; Enard W; Green RE; Burbano HA; Hublin JJ; Hänni C; Fortea J; de la Rasilla M; Bertranpetit J; Rosas A; Pääbo S (Листопад 2007). The derived FOXP2 variant of modern humans was shared with Neandertals. Curr. Biol. 17 (21): 1908–12. PMID 17949978. doi:10.1016/j.cub.2007.10.008. Загальний огляд – New York Times (2007-10-19).
- Stein, Richard A (1 жовтня 2015). Copy Number Analysis Starts to Add Up. Genetic Engineering & Biotechnology News (Paper) 35 (17): 20.
- Hopkin, Michael (16 лютого 2005). Ethiopia is top choice for cradle of Homo sapiens. Nature News. doi:10.1038/news050214-10.
- Karmin, Monika; Saag, Lauri; Vicente, Mário; Sayres, Melissa A. Wilson; Järve, Mari; Talas, Ulvi Gerst; Rootsi, Siiri; Ilumäe, Anne-Mai; Mägi, Reedik; Mitt, Mario; Pagani, Luca; Puurand, Tarmo; Faltyskova, Zuzana; Clemente, Florian; Cardona, Alexia; Metspalu, Ene; Sahakyan, Hovhannes; Yunusbayev, Bayazit; Hudjashov, Georgi; DeGiorgio, Michael; Loogväli, Eva-Liis; Eichstaedt, Christina; Eelmets, Mikk; Chaubey, Gyaneshwer; Tambets, Kristiina; Litvinov, Sergei; Mormina, Maru; Xue, Yali; Ayub, Qasim; Zoraqi, Grigor; Korneliussen, Thorfinn Sand; Akhatova, Farida; Lachance, Joseph; Tishkoff, Sarah; Momynaliev, Kuvat; Ricaut, François-Xavier; Kusuma, Pradiptajati; Razafindrazaka, Harilanto; Pierron, Denis; Cox, Murray P.; Sultana, Gazi Nurun Nahar; Willerslev, Rane; Muller, Craig; Westaway, Michael; Lambert, David; Skaro, Vedrana; Kovačevic´, Lejla; Turdikulova, Shahlo; Dalimova, Dilbar; Khusainova, Rita; Trofimova, Natalya; Akhmetova, Vita; Khidiyatova, Irina; Lichman, Daria V.; Isakova, Jainagul; Pocheshkhova, Elvira; Sabitov, Zhaxylyk; Barashkov, Nikolay A.; Nymadawa, Pagbajabyn; Mihailov, Evelin; Seng, Joseph Wee Tien; Evseeva, Irina; Migliano, Andrea Bamberg; Abdullah, Syafiq; Andriadze, George; Primorac, Dragan; Atramentova, Lubov; Utevska, Olga; Yepiskoposyan, Levon; Marjanovic´, Damir; Kushniarevich, Alena; Behar, Doron M.; Gilissen, Christian; Vissers, Lisenka; Veltman, Joris A.; Balanovska, Elena; Derenko, Miroslava; Malyarchuk, Boris; Metspalu, Andres; Fedorova, Sardana; Eriksson, Anders; Manica, Andrea; Mendez, Fernando L.; Karafet, Tatiana M.; Veeramah, Krishna R.; Bradman, Neil; Hammer, Michael F.; Osipova, Ludmila P.; Balanovsky, Oleg; Khusnutdinova, Elza K.; Johnsen, Knut; Remm, Maido; Thomas, Mark G.; Tyler-Smith, Chris; Underhill, Peter A.; Willerslev, Eske; Nielsen, Rasmus; Metspalu, Mait; Villems, Richard; Kivisild, Toomas (April 2015). A recent bottleneck of Y chromosome diversity coincides with a global change in culture. Genome Research 25 (4): 459–466. PMC 4381518. PMID 25770088. doi:10.1101/gr.186684.114.
- Schwarz, J. Uwnews.org. 17 жовтня 2007. Процитовано 10 вересня 2009.
- Richard E. Green et al; Krause, J.; Briggs, A. W.; Maricic, T.; Stenzel, U.; Kircher, M.; Patterson, N.; Li, H. та ін. (2010). A Draft Sequence of the Neandertal Genome. Science 328 (5979): 710–722. Bibcode:2010Sci...328..710G. PMID 20448178. doi:10.1126/science.1188021.
- Rincon, Paul (6 травня 2010). Neanderthal genes 'survive in us'. BBC News (BBC). Процитовано 7 травня 2010.
- Klein, Richard (1995). Anatomy, behavior, and modern human origins. Journal of World Prehistory 9: 167–198. doi:10.1007/bf02221838.
- Zimmer, Carl (21 вересня 2016). How We Got Here: DNA Points to a Single Migration From Africa. New York Times. Процитовано 22 вересня 2016.
- Bowler JM, Johnston H, Olley JM, Prescott JR, Roberts RG, Shawcross W, Spooner NA (2003). New ages for human occupation and climatic change at Lake Mungo, Australia. Nature 421 (6925): 837–40. PMID 12594511. doi:10.1038/nature01383.
- Kenneth Chang, «Neanderthals in Europe Died Out Thousands of Years Sooner Than Some Thought, Study Says», New York Times, 20 серпня 2014, процитовано 23 серпня 2014
- Beleza S, Santos AM, McEvoy B, Alves I, Martinho C, Cameron E, Shriver MD, Parra EJ, Rocha J (Січень 2013). The timing of pigmentation lightening in Europeans. Mol. Biol. Evol. 30 (1): 24–35. PMC 3525146. PMID 22923467. doi:10.1093/molbev/mss207.
- Norton HL, Kittles RA, Parra E, McKeigue P, Mao X, Cheng K, Canfield VA, Bradley DG, McEvoy B, Shriver MD (Березень 2007). Genetic evidence for the convergent evolution of light skin in Europeans and East Asians. Mol. Biol. Evol. 24 (3): 710–22. PMID 17182896. doi:10.1093/molbev/msl203. Загальний огляд – Science Magazine.
{kind=link}
Посилання
- Palaeos
- Хронологія гомінідів
- Еволюція Берклі
- Історія еволюції тварин
- Tree of Life Web Project — інтерактивно дослідити повне філогенетичне дерево
- Хронологія людини (Інтерактивна) — Смітсонівський інститут, Національний музей природознавства (серпень 2016).